
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Рефераты по косметологии
Рефераты по криминалистике
Рефераты по криминологии
Рефераты по науке и технике
Рефераты по кулинарии
Рефераты по культурологии
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Рефераты по косметологии
Рефераты по криминалистике
Рефераты по криминологии
Рефераты по науке и технике
Рефераты по кулинарии
Рефераты по культурологии
Реферат: Біофізичні основи електрографії
Реферат: Біофізичні основи електрографії
Біофізичні основи електрографії
1. Поняття електрографії
Робота багатьох важливих органів людського організму зв'язана з електричними явищами, що виникають унаслідок генерації потенціалів дії на мембранах м'язових і нервових кліток. При цьому в різних тканинах організму відбувається зміна згодом електричних потенціалів. Електрична активність кліток, тканин, органів зв'язана з їхнім функціональним станом. Реєстрація різностей потенціалів між різними крапками організму лежить в основі електрографічних діагностичних методик, з яких найбільш відомою і поширеною є методика електрокардіографії (ЕКГ).
Вивчення біофізичних основ електрографії вимагає попереднього поглиблення знань в області фізики електромагнітних явищ.
Електричне поле -
це вид матерії, що породжується електрично зарядженими тілами чи перемінним
магнітним полем і виявляється по дії на заряджені тіла. Характеристикою
зарядженого тіла є його заряд. Заряд тіла - це скалярна фізична величина. Від
значення заряду тіла залежить величина електромагнітних взаємодій, у яких бере
участь це тіло. Найважливішими характеристиками електричного поля є його
напруженість ( ![]() ) і потенціал (
) і потенціал (![]() ).
).
Напруженість (![]() ) - це
векторна фізична величина, що є силовою характеристикою електричного поля.
Напруженість у деякій крапці електричного поля дорівнює відношенню сили (
) - це
векторна фізична величина, що є силовою характеристикою електричного поля.
Напруженість у деякій крапці електричного поля дорівнює відношенню сили (![]() ), що діє з
боку поля на крапкове заряджене тіло, поміщене в цю крапку, до його заряду (q),
тобто
), що діє з
боку поля на крапкове заряджене тіло, поміщене в цю крапку, до його заряду (q),
тобто
![]() =
=![]()
![]() (1)
(1)
У СИ напруженість поля виміряється у вольтах на метр (В/м).
Потенціал (![]() ) - це
скалярна величина, що є енергетичною характеристикою електричного поля.
Потенціал поля в деякій крапці дорівнює відношенню потенційної енергії (W),
якою володіє крапкове заряджене тіло, поміщене в цю крапку, до його заряду,
тобто
) - це
скалярна величина, що є енергетичною характеристикою електричного поля.
Потенціал поля в деякій крапці дорівнює відношенню потенційної енергії (W),
якою володіє крапкове заряджене тіло, поміщене в цю крапку, до його заряду,
тобто
![]() (2)
(2)
Потенціал, як і
напруга (![]() різниця потенціалів), у СИ виміряється у вольтах (В ).
різниця потенціалів), у СИ виміряється у вольтах (В ).
Напруженість зв'язана з потенціалом співвідношенням
E = -grad(![]() )
)
Таким чином,
напрямок вектора![]() збігається з напрямком
найбільшого убування потенціалу.
збігається з напрямком
найбільшого убування потенціалу.
Будемо тут і далі називати крапковим зарядом заряджене тіло, розмірами якого можна зневажити. Напруженість електричного поля, створюваного в деякій крапці відокремленим крапковим зарядом q, обчислюється по формулі
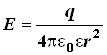 (3)
(3)
де ![]() - діелектрична проникність середовища;
- діелектрична проникність середовища; ![]() о – електрична постійна (
о – електрична постійна (![]() 0 = 8,85 х 10-12 Ф/м), а м -
відстань від крапки, у якій визначається напруженість поля, до крапки, у якій
знаходиться заряд.
0 = 8,85 х 10-12 Ф/м), а м -
відстань від крапки, у якій визначається напруженість поля, до крапки, у якій
знаходиться заряд.
Потенціал поля, створюваного крапковим зарядом, розраховується по формулі
![]() . (4)
. (4)
Якщо поле створюється декількома зарядами, то його характеристики (напруженість і потенціал) розраховуються, виходячи з принципу суперпозиції полів. Відповідно до цього принципу, напруженість електричного поля, створеного в деякій крапці декількома зарядами, дорівнює векторній сумі напруженості полій, створених у цій крапці кожним із зарядів, а потенціал - алгебраїчній сумі потенціалів, створених кожним із зарядів, тобто
![]() =
= ![]() 1+
1+![]() 2 +
2 +![]() 3.....+
3.....+
![]() n;
(5)
n;
(5)
![]() , (6)
, (6)
де ![]() і
і ![]() - результуючі
напруженості і потенціалу поля, створеного n зарядами;
- результуючі
напруженості і потенціалу поля, створеного n зарядами; ![]() 1,
1, ![]() 2,
2, ![]() n -
напруженості полів, створених кожним із зарядів, q1, q2,
... , qn - потенціали полів, створених кожним із зарядів.
n -
напруженості полів, створених кожним із зарядів, q1, q2,
... , qn - потенціали полів, створених кожним із зарядів.
Виходячи з
принципу суперпозиції полів, можна розрахувати характеристики поля створюваного
різними системами зарядів. Найпростішою системою зарядів є електричний диполь -
система, що складається з двох крапкових однакових по модулі, але протилежних
за знаком зарядів (+ q і -q), що знаходяться на відстані L друг від друга.
Відстань L називається плечем диполя. Характеристикою електричного диполя є
дипольний момент. Дипольний момент (![]() ) електричного диполя - це
векторна величина, рівна по модулі добутку модуля одного з зарядів, що утворять
диполь, на плече диполя, і спрямована від негативного заряду диполя до
позитивного
) електричного диполя - це
векторна величина, рівна по модулі добутку модуля одного з зарядів, що утворять
диполь, на плече диполя, і спрямована від негативного заряду диполя до
позитивного
![]() =q
=q![]() (7)
(7)
При цьому плече
диполя![]() розглядається як вектор, спрямований від негативного заряду до позитивного.
розглядається як вектор, спрямований від негативного заряду до позитивного.
Використовуючи принцип суперпозиції полів, можна показати, що потенціал поля, створеного диполем у крапці, розташованої на відстані r від центра диполя, у випадку, якщо r » L, розраховується по формулі.
![]() , (8)
, (8)
де ![]() - кут між прямою,
проведеної від центра диполя до крапки, у якій визначається потенціал (крапка А
на рис. 1), і вектором дипольного моменту.
- кут між прямою,
проведеної від центра диполя до крапки, у якій визначається потенціал (крапка А
на рис. 1), і вектором дипольного моменту.
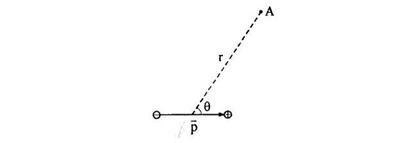
Рис.1.
Електричні явища в організмі людини зв'язані з протіканням електричних струмів у тканинах організму, що є здебільшого середовищами, що добре проводять електричний струм. Основними характеристиками струмів є сила (I) і щільність струму (j ). Якщо через провідник протікає струм, то сила струму – це відношення заряду (q), що проходить через поперечний переріз провідника за деякий проміжок часу (t), до цього проміжку, тобто
I=![]() (9)
(9)
У СИ сила струму
виміряється в амперах ( А).![]()
Щільність струму – це відношення сили струму, що протікає через поперечний переріз провідника, до площі цього перетину (S),
j = ![]() (10)
(10)
Щільність струму виміряється в амперах на метр квадратний (А/м2).
Здатність провідника проводити струм характеризується його опором (R).
Опір провідника ![]() виміряється в
омах (Ом). Опір провідника залежить від його геометричних розмірів і від
провідних властивостей матеріалу, з якого зроблений провідник. Ці провідні
властивості матеріалу (середовища) можна охарактеризувати такими величинами, як
питомий опір і питома електропровідність (питома провідність).
виміряється в
омах (Ом). Опір провідника залежить від його геометричних розмірів і від
провідних властивостей матеріалу, з якого зроблений провідник. Ці провідні
властивості матеріалу (середовища) можна охарактеризувати такими величинами, як
питомий опір і питома електропровідність (питома провідність).
Питома електропровідність речовини (σ) – це величина, зворотна його питомому опору.
От значення питомої електропровідності (Ом-1м-1) різних біологічних тканин : спинномозкова рідина – 1,8; кров – 0,6; м'язова тканина – 0,5; нервова тканина – 7х10-2 ; жирування тканина – 3х10-2; суха шкіра – 10-5; кістка без окістя – 10-7.
Величини струмів, що протікають у середовищах, залежать від провідних властивостей цих середовищ і від напруженості електричного поля в них. Зв'язок між цими величинами встановлює закон Ома в диференціальній формі
![]() (11)
(11)
Найпростішою моделлю токових систем є відокремлене крапкове джерело струму. Воно характеризується силою струму, що випливає з нього (чи втікає в нього). Якщо струм випливає з крапкового джерела струму, то таке джерело називається джерелом, якщо втікає – стоком. У першому випадку струм джерела приймається рівним + I, у другому - -I.
Залежність
напруженості поля, створюваного відокремленим крапковим джерелом струму, від r така ж, як і в
напруженості поля, створюваного відокремленим крапковим зарядом. Можна
показати, що і для інших токових систем формули для напруженості і потенціалів
створюваних полів подібні таким в аналогічних систем зарядів. Різниця полягає в
тому, що у формулах для напруженості і потенціалів полів токових систем стоять
відповідно величини I і σ замість величин q і![]() у формулах для напруженості і
потенціалів полів систем заряджених тіл відповідно.
у формулах для напруженості і
потенціалів полів систем заряджених тіл відповідно.
Розглянемо токовий диполь, тобто систему з двох крапкових джерел струму (джерела і стоку), сили струмів яких рівні по модулі. Характеристиками такого диполя є сила струму крапкових джерел, плече диполя (відстань між джерелами), що, як і у випадку електричного диполя, позначається L, момент токового диполя (дипольний момент), модуль якого (D) обчислюється по формулі
D =IL. (12)
Момент токового диполя – це векторна величина, спрямована від стоку до джерела.
Потенціал поля, створюваного довільною системою струмів, може бути представлений у виді ряду
![]() (13)
(13)
А, В, С, D, ... - деякі коефіцієнти. Таке представлення потенціалу поля, створеного системою струмів, називається мультипольним розкладанням.
Якщо система струмів скомпенсована, тобто сила струмів, що випливають із джерел, дорівнює силі струмів, що втікають у стоки, то у формулі (13) А = 0 і ця формула здобуває вид
![]() (14)
(14)
Зі збільшенням r кожний з наступних членів цього ряду убуває швидше, ніж попередній, тому
при досить великих r значимим виявляється тільки перший доданок
цього ряду ![]() .
З іншого боку, саме таку залежність потенціалу поля від r має токовий диполь. Звідси випливає, що якщо потенціал поля,
створюваного складною скомпенсованою системою струмів, визначається на
відстанях, значно більших r, від цієї системи, то систему можна з хорошою
точністю вважати токовим диполем.
.
З іншого боку, саме таку залежність потенціалу поля від r має токовий диполь. Звідси випливає, що якщо потенціал поля,
створюваного складною скомпенсованою системою струмів, визначається на
відстанях, значно більших r, від цієї системи, то систему можна з хорошою
точністю вважати токовим диполем.
Більшість процесів, що протікають в організмі, зв'язано зі зміною електричних потенціалів клітинних мембран. Так, у м'язовій клітці цей процес викликає її скорочення, у нервовій клітці поширення по мембрані хвилі потенціалу дії являє собою передачу нервового імпульсу. В усіх випадках, коли говориться про потенціали дії, мається на увазі різниця потенціалів між збудженим і, що знаходяться в спокої ділянками збудливої тканини.
2. Нервові імпульси
Процес нервового порушення, що поширюється, є коливальним, переривчастим процесом. Це встановлено шляхом вивчення електричних явищ, що відбуваються в нерві при його порушенні. При порушенні нервових волокон (і нервових кліток) їхня збуджена ділянка стає електронегативною стосовно ще спочиваючих ділянок унаслідок появи негативно заряджених іонів на зовнішній поверхні нервового волокна в ділянці, охопленій порушенням. Тому, з'єднуючи з осцилографом — приладом для реєстрації слабких електричних коливань, що швидко протікають — дві крапки нерва, одна з яких у даний момент охоплена процесом збудження, а інша знаходиться в стані спокою, можна записати так називані струми, чи потенціали, дії, що виникають при зміні електричних зарядів нерва (рис. 2).
| Распр |
|
|
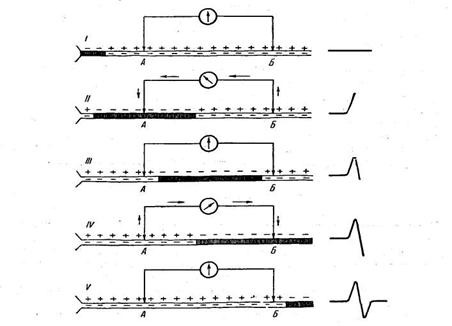
Рис.2. Схема виникнення струмів дії при поширенні нервового імпульсу по нервовому волокну.
Область нервового волокна, охоплена в даний момент збудженням, заштрихована. Знаки + і — показують розподіл позитивно і негативно заряджених іонів усередині волокна і на його зовнішній поверхні. Ділянки А и В приєднані до осцилографу (кружок зі стрілкою, що показує напрямок струму).
I - збудження, що біжить по нервовому волокну від тіла нервової клітки, ще не дійшло до А; різниці потенціалів між Л и Б тому немає. II - збудження охопило область А; ділянка А електронегативна у відношенні Б; дзеркальце осцилографа відхиляється, що реєструється в записі струму дії у виді його висхідного коліна (права сторона рисунка). III- збудження, що поширюється по нерві, пішло з ділянки А, але ще не дійшло до ділянки Б- потенціал цих ділянок тому знову однаковий і дзеркальце осцилографа повертається в нульове положення. IV - збудження дійшло до ділянки Б, що є тепер електронегативним стосовно ділянки А (тому осцилограф реєструє виникнення струму, спрямованого в зворотну сторону в порівнянні з його напрямком у записі II).
V- збудження вийшло за межі з'єднаних з осцилографом ділянок нерва і різниця потенціалів між ділянками А и Б знову зникає.
Кожна ділянка нерва при своєму порушенні здобуває негативний заряд, причому ця електронегативність наростає, досягає максимуму і потім зникає протягом тисячних (а в деяких видах нервових волокон ссавців протягом десятитысячних) часток секунди. Слідом за цим, якщо збудження продовжується, знову розвивається електронегативність, знов-таки настільки ж швидко зникаюча. Таким чином, при нанесенні на нерв ряду повторних роздратувань і навіть при нанесенні безупинно діючого роздратування в кожній крапці нервового волокна спостерігається ряд наступних друг за другом коливань електричного потенціалу. Ні посилення, ні частішання, ні подовження роздратувань не усувають переривчастості струмів, що відводяться від нерва. Отже, у нервовій тканині при її порушенні відбувається ряд надзвичайно швидких і легко оборотних реакцій, що обумовлюють короткочасне нагромадження на поверхні збудженої ділянки неврона надлишок негативно заряджених іонів. Короткочасне, швидко (за соті, тисячні чи десятитисячні частки секунди) наростаюче і спадаюче порушення, що поширюється по неврону і характеризується зміною фізико-хімічних властивостей і електричного потенціалу збудженого в даний момент ділянки неврона, називається нервовим імпульсом. Кожен нервовий імпульс характеризується як би окремим циклом хімічних і фізико-хімічних змін, що поширюються по неврону й обумовлюють виникнення різниці потенціалів між ділянкою неврона, що знаходиться в даний момент у стані порушення, і його ще не збудженими (чи уже вихідними зі стану порушення) ділянками.
Ця різниця потенціалів позначається як струм дії, потенціал дії, чи біострум (ці терміни застосовуються також для позначення різниці потенціалів, що виникає при порушенні інших тканин).
Дослідження струмів дії в даний час є дуже важливим засобом вивчення явищ, що відбуваються в нервовій системі при передачі імпульсів порушення, тому що воно дозволяє безпосередньо вивчати зміни, що відбуваються в нервовій тканині за надзвичайно короткі інтервали часу — тисячні частки секунди. Електричні зміни в тканинах енергетично виражаються незначною величиною, складаючи, наприклад, у м'язі менш 0,00001 частини всієї кількості теплової і механічної енергії, що звільняється при скороченні. Однак високий розвиток електровимірювальної і підсилювальної техніки робить електрофізіологічну методику дослідження однієї з найбільш зроблених і точних при вивченні динаміки порушення в нервовій і м'язовій системах. Початок вивченню електричних явищ у тваринних тканинах було покладено в 1786 р. відкриттям болонського лікаря і фізіолога Гальвані. Гальвані помітив, що якщо між нервом і м'язом отпрепарованої лапки жаби встановлюється замикання за допомогою металевого провідника, то м'яз здригається, як при пропущенні через неї розряду лейденської банки. Гальвані витлумачив цей факт як прояв «тваринної електрики». Він вважав, що нерви і м'язи заряджені протилежною електрикою і що металевий провідник у цьому досвіді тільки замикає ланцюг між нервом і м'язом.
Сучасник Гальвані - фізик Вольта, професор університету в Павії, витлумачив результати досвідів Гальвані як наслідок того, що металевий провідник у його досвідах був неоднорідним. При зіткненні металу з вологою тканиною виникала різниця потенціалів між металом і рідиною, причому внаслідок неоднорідності металевого провідника на його кінцях, що стикалися з тканиною, створювалися неоднакові заряди, що приводило до виникнення електричного струму.
Обоє дослідника виявилися праві кожен по-своєму. Гальвані знайшов варіант досвіду, у якому скорочення отпрепарованого м'яза досягалося без участі металевих провідників у результаті простого накидання на м'яз її ж нерва (так називаний досвід зі скороченням без металів). Цим досвідом незаперечно доводилося, що джерелом електрики може бути сама збудлива тканина, і тим самим відкривалася нова велика глава фізіології, що одержала згодом назву електрофізіології.
Вольта ж прийшов до відкриття першого джерела постійного струму («вольтова стовпа»), у якому різниця потенціалів, чи, по термінології Вольта, електрорушійна сила, виходила при взаємодії металу з рідиною. Вольтов стовп складався спочатку з багатьох пар мідних і цинкових кружків, накладених один на одного і розділених сукняними прошарками, змоченими солоною водою. (Найбільш могутня для того часу батарея, що складалася з 4200 пар і цинкових пластинок, була створена в 1802 р. російським академіком В. В. Петровим, що, поряд з фізичними дослідженнями, привели його до відкриття електричної дуги, ставив також досвіди по впливі електричного струму на водяних тваринах.).Цим відкриттям був покладений початок навчанню про електричний («гальванічний») струм і були спростовані віталістичні представлення про специфічну природу «тваринної електрики».
Міцні основи експериментального вивчення біоелектричних властивостей м'яза і нерва були закладені пізніше, починаючи із середини минулого століття, особливо дослідженнями Дюбуа-Реймона, Германа, И. М. Сєченова, Н. Е. Введенського, що застосували для дослідження біострумів їхню реєстрацію за допомогою фізичних приладів — гальванометрів. Особливо плідним у цьому періоді розвитку електрофізіології було застосування Н. Е. Введенським телефонічної методики.
3. Струми спокою м'яза і нерва
Якщо подовжню поверхню м'яза чи нерва і поверхню нанесеного на них поперечного розрізу з'єднати електродами, що не поляризуються з чуттєвим гальванометром, то гальванометр знайде наявність струму, спрямованого в зовнішньому ланцюзі від подовжньої поверхні м'яза чи нерва до поперечного їх перетину. Іншими словами, поперечний переріз тканини виявляється електронегативним стосовно її подовжньої поверхні. Струм, що спостерігається при з'єднанні подовжньої і поперечної поверхонь тканини, Дюбуа-Реймон назвав струмом спокою.
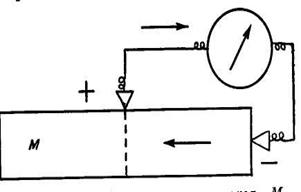
Рис.3. Схема відведення струму спокою. М - м'яз (чи нерв). Поперечний розріз м'яза (нерва) заряджений електронегативно (-) стосовно неушкодженої поверхні (+). Стрілки - напрямок струму у внутрішньому і зовнішньому ланцюзі.
Однак Герман показав, що так званий струм спокою виникає лише в результаті ушкодження тканини і причиною його є різниця потенціалів між неушкодженою частиною тканини (подовжньою поверхнею) і ушкодженою її ділянкою, яким є поперечний розріз. Таким чином, струм спокою не передує у нерві чи м'язі, як вважав Дюбуа-Реймон, а виникає при ушкодженні тканини і взагалі при місцевих змінах (альтераціях) тканини під впливом різних фізіологічних і хімічних впливів. Тому він одержав назву струму ушкодження, чи альтераційного струму.
Виявилося також, що досвід Гальвані зі скороченням без металів не вдається, якщо м'яз отпрепарован дуже ретельно, без ушкоджень. Але якщо м'яз попередньо поранити, то накидання на неї нерва веде незмінно до скорочення, якщо нерв торкається як пораненої, так і неушкодженої частини.
Найбільша різниця потенціалів, що доходить на м'язі до 0,08 V, спостерігається слідом за нанесенням ушкодження. Протягом першої години струм ушкодження падає майже до половини своєї вихідної величини. У нерві виникаюча при ушкодженні різниця потенціалів звичайно не перевищує, 0,03 V і з часом убуває швидше, ніж у м'язі. У нерві людини вона складає лише близько 0,006 V, що зв'язано, мабуть, з достатком з'єднанних прошарків, що є побічним замиканням для струму, що відводиться від нерва. У процесі дегенерації нерва величина виникаючого при ушкодженні струму перетерплює поступове зменшення, і через 10 днів у теплокровних чи через 30 днів у холоднокровних тварин він уже не виявляється.
Перший прямий доказ переривчастої, коливальної природи порушення в нерві дане Н. Е. Введенським (1883), що примінив телефон для вислуховування струмів дії нерва. Відведення двох крапок нерва на телефон дозволило йому вислухувати ритми діяльності нерва при роздратуваннях різної сили і частоти. Користаючись цією методикою, Н. Е. Введенський ще в 80-х роках минулого століття зробив ряд основних висновків про ритми порушення; ці висновки надалі були підтверджені іншими дослідниками в умовах графічної реєстрації.
Для графічної реєстрації струмів дії, що звичайно представляються у виді швидких електричних коливань, пізніше були застосовані спеціальні гальванометри чи електрометри, що володіють дуже малою інерцією системи, що реєструє. Такими є капілярний електрометр, струнний гальванометр і осцилографи.
Капілярний електрометр у своїй істотній частині представляє капілярну скляну трубку, що містить ртуть зі слабким розчином сірчаної, кислоти що знаходиться, над нею. На поверхні зіткнення ртуті з кислотою виникає різниця потенціалів, причому ртуть заряджається позитивно, а кислота негативно. Наявність заряду зменшує поверхневий натяг ртутного меніска унаслідок взаємного відштовхування однойменно заряджених часток ртуті. Якщо через цю систему пропустити струм, наприклад, у такому напрямку, щоб меніск з'явився катодом, а сірчана кислота — анодом, то заряд меніска зменшиться. Завдяки цьому, поверхневий натяг ртуті збільшиться і меніск опуститься, змістившись в напрямку струму, що пропускається. При зворотному напрямку струму збільшення заряду ртутного меніска зменшить його поверхневий натяг і меніск підніметься. Таким чином, зсув ртутного меніска відбуваються завжди в напрямку діючого струму. Рух ртутного меніска можна спостерігати за допомогою мікроскопа чи фотографувати на світлочутливий папір.
Інерція ртутного меніска усе-таки відносно велика, і більш досконалим приладом є струнний гальванометр. Дуже тонка провідна нитка цього гальванометра, натягнута вертикально між полюсами електромагніта, може відтворювати без значних перекручувань до 500 коливань у секунду.
Значно більшу частоту коливань можуть відтворювати без перекручування шлейфні, чи магніто-електричні, осцилографи, що знайшли собі електрофізіологічне застосування. Рухливою системою тут є шлейф - петля з металевої стрічки, перекинута від одного затиску до іншого через блок, що відтягається догори пружиною. Петля міститься між полюсами постійного магніту. При проходженні струму обидві половини петлі відхиляються в протилежні сторони, тому що напрямок струму в них по-різному. Унаслідок цього вся петля повертається на деякий кут, величина якого визначається силою підводимого струму. Відповідно зміщається пучок світла, відбиваний укріпленим на петлі дзеркальцем. Відхилення світлового пучка фотографуються. Такі осцилографи можуть відтворювати без перекручування до 10 000 коливань у секунду.
Однак безпосереднє використання шлейфних і катодних осцилографів для реєстрації струмів дії є неможливим унаслідок недостатньої чутливості цих приладів. Тому струми дії, що підводяться до осцилографу, необхідно значно підсилювати. Це досягається за допомогою підсилювачів.
Струм дії
порозумівається, як уже було зазначено, тим, що збуджена ділянка тканини
виявляється на короткий час електронегативною стосовно
спочиваючих ділянок. Коли один з електродів, що
відводять, прикладений до неушкодженої, а інший - до ушкодженої
ділянки нерва чи м'яза, різниця потенціалів між електродами при проходженні
хвилі порушення під першим електродом на короткий час
зменшиться: прилад, що реєструє, знайде коливання, що швидко протікає в
напрямку, зворотному напрямку струму спокою. Що ж стосується ушкодженої ділянки
(під другим електродом), то хвиля збудження, що приходить вгасає в ньому, не
змінюючи його електричного стану. У цьому випадку струм дії має форму
однофазного, тобто однобічного коливання (рис.4, а).
Якщо ж обидва електроди розташувати на неушкодженій подовжній поверхні препарату, то струм дії представиться у виді двофазного коливання (рис.4, б), тому що ділянки тканини, на яких лежать електроди, з'єднані з гальванометром, не одночасно охоплюються порушенням. Поширюючись з визначеною швидкістю, збудження спочатку доходить до найближчого електрода і створює тут швидко минаючу електронегативність; прилад, що реєструє, дає коливання в одному напрямку. Коли ж при подальшому своєму поширенні порушення, минавши область першого електрода, досягає ділянки, що лежить під другим електродом, у зовнішньому ланцюзі виникає короткочасний струм зворотного напрямку, що регіструється у виді другого коливання, спрямованого в протилежну сторону.
|
|

Рис.4. Ліворуч однофазний, праворуч двофазний струм дії сідничного нерва жаби, зареєстрований на катодному омосцилографі.
Тривалість однофазного струму дії в кістякових м'язах хребетних тварин виражається тисячними частками секунди: у м'язах жаби вона складає 0,003-0,004 секунди, у м'язах теплокровних - 0,002-0,003 секунди. У нервах тривалість струму дії складає 0,0005-0,0020 секунди. Ніж скоріше проводиться порушення в нерві, тим швидше протікає струм дії.
4. Походження струмів спокою і струмів дії
Ще Сєченов відзначав значення обміну речовин у виникненні різниці потенціалів у тваринних тканинах. У розвиток цих поглядів В. Ю. Чаговец (1896) показав можливість пояснення електричних явищ у м'язах і нервах, виходячи з представлень про концентраційні струми, обумовлених різною концентрацією деяких іонів (наприклад, Н-іонів) у різних ділянках тканини. Різниця концентрацій може бути результатом зміни обмінних процесів у ділянках збудження чи ушкодження.
Пізніше одержала поширення мембранна теорія (Ю. Бернштейн, 1902), заснована на схематичних представленнях про існування на поверхні м'язових і нервових волокон особливого прикордонного шару (мембрани), що, подібно деяким штучно одержуваним перетинкам, є прохідною для катіонів і непрохідною для аніонів. Тому що концентрація електролітів по обох сторонах мембрани неоднакова, то катіони (наприклад, що містяться у відносному надлишку усередині волокна іони калію) прагнуть перейти в зовнішнє середовище, але утримуються на зовнішній поверхні мембрани зарядами своїх аніонів. Унаслідок цього мембрана стає поляризованою: зовнішня поверхня її заряджена позитивно, а внутрішня – негативно( рис 5.). Поки волокно не ушкоджене, по всій його поверхні потенціал однаковий, але при його ушкодженні, тобто при порушенні цілості мембрани, негативні іони одержують можливість дифундувати зсередини назовні: ушкоджене місце виявляється електронегативним стосовно нормально поляризованої поверхні.
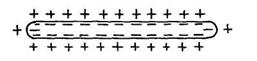
Рис.5. Схема неушкодженого поляризованого нервового волокна (у стані спокою).
Таке походження струму спокою. Збудження ж зв'язане зі збільшенням проникності мембран, про що свідчить ряд спостережень. Завдяки цьому оболонка в ділянці порушення стає однаково проникною для всіх іонів і деполяризується, унаслідок чого збуджена ділянка виявляється електронегативною стосовно іншій позитивно зарядженій поверхні волокна. На відміну від деполяризації, що створюється ушкодженням, деполяризація, зв'язана з порушенням, є оборотною і поширюється разом зі збудженням. Саме поширення збудження з цього погляду представляється, як поширення плани деполяризації. На границі між деполяризованою і поляризованою ділянкою виникає внаслідок різниці потенціалів переміщення іонів, що приводить до деполяризації нової ділянки і до відновлення поляризації колишнього (рис.6).
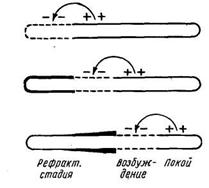
Рис.6. Схема, що показує деполяризацію мембрани нервового волокна в ділянці збудження (ця ділянка показана переривчастою лінією); збудження поширюється ліворуч праворуч і залишає за собою рефракторну фазу (охоплена нею ділянка обкреслена жирною лінією). Стрілки показують виникнення місцевого струму.