
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Рефераты по косметологии
Рефераты по криминалистике
Рефераты по криминологии
Рефераты по науке и технике
Рефераты по кулинарии
Рефераты по культурологии
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Рефераты по косметологии
Рефераты по криминалистике
Рефераты по криминологии
Рефераты по науке и технике
Рефераты по кулинарии
Рефераты по культурологии
Книга: Ихтиология
Книга: Ихтиология
Ихтиология
Анисимова И.М., Лавровский В.В.
Из-во "Высшая школа"
1983 г.
Часть I. Общая ихтиология. Место рыб в системе животных
· Рыбы (и круглоротые) в системе животных занимают самое низкое место среди позвоночных. Они относятся к типу хордовых Chordata (благодаря наличию хорды–эластичного тяжа, являющегося у них начальным осевым скелетом, у большинства рыб замещающегося позвоночником), подтипу позвоночных или черепных Vertebrata или Craniata, надклассу рыбы Pisces. В этом надклассе среди современных рыб выделяются два класса – хрящевые рыбы Chondrichthyes и костные рыбы Osteichthyes. К подтипу позвоночных относится и класс круглоротых Cyclostomata – миноги и миксины; менее совершенные, чем настоящие рыбы, они не имеют ещё челюстей и парных плавников и называются рыбообразными. Положение основных групп рыб в системе животных показано на рис. 1.
·
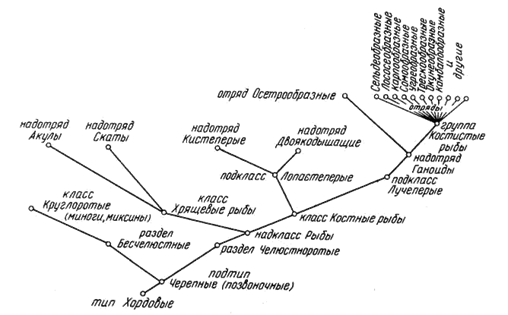
Рис. 1. Положение основных групп рыб в системе животных (по Наумову, Карташеву, 1979, с сокращениями)
· Рыбы – древняя группа, насчитывающая сотни миллионов лет. Наиболее ранние ископаемые остатки позвоночных принадлежат круглоротым и панцирным рыбам, найденным в Европе и Америке в силурийских отложениях (палеозойская эра). Примитивные рыбообразные животные по строению были близки к круглоротым, они появились и обитали в пресных водах.
· Позднее возникли акулы и скаты. Их появление относят к концу палеозойской эры (карбонский и пермский периоды) и к началу мезозойской эры (триасовый и юрский периоды), но затем эта группа начала угасать.
· К палеозойской эре приурочивают исследователи и появление относительно сложноорганизованных рыб, характеризующихся более или менее окостеневшим скелетом. В среднем девоне древние лучёперые, кистепёрые и двоякодышащие рыбы уже многочисленны в пресных водах, а в мезозойской эре начинается переход лучепёрых в моря.
· Костистые рыбы Teleostei, обнаруженные в мезозойской эре (триасовый и меловой периоды), господствовали как в пресных, так и в морских водоемах.
· В настоящее время насчитывают более 20 тыс. видов рыб, живущих в морских и пресных водах. В водах СССР встречается около 1500 видов, из них около 300 видов пресноводные рыбы, остальные – морские.
· Биологические группы рыб. В соответствии с зоной обитания в водоемах различных типов выделены следующие биологические группы рыб: морские рыбы – живут только в соленой воде морей и океанов (пеламида, тунец, скумбрия, анчоус и др., всего около 11,6 тыс. видов); пресноводные рыбы – обитают только в пресных водах (карась, щука и др. , всего около 8,3 тыс. видов); солоновато водные рыбы – живут в солоноватой воде опресненных участков морей, предустьевых пространств (бычки, речная камбала и др. ); проходные рыбы – в определённые периоды жизни меняют морскую среду на пресноводную или наоборот; при этом морские заходят для нереста в реки, до их верховьев, а пресноводные выходят из рек нереститься в море (осетр, белуга, лососевые рода Oncorhynchus и др. , всего около 130 видов); полупроходные рыбы – это обитатели опресненных пространств морей, поднимающиеся на нерест невысоко в реки (сазан, лещ, вобла, сом, судак).
· По приуроченности к характерным экологическим зонам водоёма – пелагиали (толща воды), бентали (придонная зона), литорали (прибрежная зона) – различают рыб пелагических, бентических, литоральных.
· В пределах этих больших групп выделены более узкие экологические группы в связи с особенностями питания, размножения и т. д.
Некоторые абиотические факторы и их влияние на рыб
· Известно, что свыше 71 % поверхности земного шара (~360 млн. км2 из510) занимают моря, океаны и внутренние водоемы. Максимальная известная глубина океана – примерно 11 000 м, а около 50–60% площади океанов приходится на глубины 3000м. Высота некоторых горных озер превышает 6000 м над уровнем моря. Рыбы распространены повсюду. Они обитают от полярных вод до тропиков, от поверхностных слоев до океанических глубин, в пресной и морской воде, у берегов и в открытых пространствах, у дна и в толще воды, в высокогорных и пещерных водоемах, в горячих источниках и при отрицательной температуре, в условиях заморных водоемов.
· Очевидно, в процессе эволюции у них выработались как общие, так и весьма специфические приспособления, позволяющие всем им, с одной стороны, жить в водной среде, а с другой – осваивать столь различные водоёмы. Эти приспособления затрагивают все системы организма. К общим, наиболее характерным признакам рыб относятся жабры, дающие возможность использовать для дыхания растворенный в воде кислород; форма тела и плавники, обеспечивающие передвижение в воде; плавательный пузырь, выполняющий гидростатическую функцию; метамерное строение двигательной мускулатуры, благодаря чему в движении участвует почти все тело; кожа, в которой образуется чешуя, а также слизь, играющие важную роль в защите тела от внешних воздействий и во многих других процессах; боковая линия – важный сейсмосенсорный орган; многочисленные приспособления, обеспечивающие сохранение потомства при наружном осеменении икры и эмбриональном развитии вне материнского организма.
· Приспособлением к жизни в водной среде (в условиях пониженной температуры, низкого содержания кислорода) явился и низкий уровень обмена. По способности приспосабливаться к окружающей среде рыбы превосходят всех других позвоночных.
· Рыбы – первичноводные животные, всю жизнь проводящие в воде, поэтому свойства воды оказывают сильнейшее влияние на жизнедеятельность и в конечном счете на состав ихтиофауны водоема.
· Плотность воды и движение рыб. Высокая плотность и малая вязкость воды (определяемые количеством растворенных в ней солей и температурой) играют большую роль в жизни рыб. С этими факторами связаны, с одной стороны, сопротивление воды движущемуся телу, а следовательно, и скорость его передвижения, а с другой давление массы воды на организм, которое, увеличиваясь с глубиной на каждые ~10м на 1,01 • 105Па, в глубинах океана превышает 1013 • 105 Па.
· Поэтому огромное значение для рыб имеют приспособления, обеспечивающие существование в толще воды, удержание тела в определённом положении и облегчение плавания.
· Эти приспособления выразились в снижении плотности тела, благодаря чему увеличивается способность держаться в толще воды без особых усилий, и в выработке разнообразных форм тела и способов движения.
· Плотность тела рыб только немного выше этих показателей воды, а у хороших пловцов – равна им. Благодаря этому плавучесть (отношение плотности тела рыбы к плотности воды) становится нулевой или нейтральной (осетр, судак, карп, голавль и другие нектонные рыбы). Таким образом, рыбы в воде относительно невесомы. У придонных и донных рыб отмечена незначительная отрицательная плавучесть (у камбал и бычков – 0,05–0,07) – это способствует удержанию их у дна.
· Выравнивание плотности тела рыбы и воды происходит у хрящевых и некоторых костистых рыб за счет накопления жира. Костистые рыбы имеют специальный гидростатический аппарат – плавательный пузырь –мешкообразный орган, наполненный газами. Изменение объёма плавательного пузыря позволяет рыбе в известных пределах регулировать плавучесть. Специальные гидростатические приспособления имеют рыбы и в эмбриональном периоде: оводнение желтка, наличие в желтке жировых капель, увеличение перивителлинового пространства в икринках и др.
· Термика водоема. Исключительно важную роль имеют свойства воды, определяющие термический режим водоема. Постепенность нагрева и охлаждения водных масс обеспечивают: очень высокая удельная теплота парообразования (2255 Дж/г); высокая скрытая теплота плавления воды (334 Дж/г), из-за которой большое количество тепла выделяется при ее замерзании и поглощается при таянии льда; более высокая, чем у суши, теплопрозрачность воды, которая способствует проникновению солнечных лучей в глубину и непосредственному нагреванию толщи воды на разных уровнях; очень высокая теплоемкость воды (4 Дж/г), т. е. способность поглощать теплоту. Низкая теплопроводность льда ведет к тому, что зимой температура воды с глубиной увеличивается и держится в придонных слоях водоема на уровне нескольких градусов, а снижение удельного веса воды при замерзании, ее способность при этом увеличиваться в объёме приводит к всплыванию льда, который образует на поверхности теплоизоляционный слой. Все это исключает промерзание водоемов, кроме совсем мелководных.
· Основную массу тепловых лучей из атмосферы поглощают верхние слои воды. Количество лучей, проникающих вглубь, постепенно убывает, поэтому температура воды на разных глубинах неодинакова (температурная стратификация). Зимой в малых водоемах наблюдается обратная стратификация – от 0°С под ледовым покровом к 4°С на глубинах.
· Рыбы относятся к пойкилотермным животным, т, е. к животным с переменной температурой тела (от слова “пойкилос” – пестрый), или, как неудачно их называют, к холоднокровным животным.
· Тепло, вырабатываемое в организме рыб в обменных процессах, не задерживается в теле, так как у них нет механизмов, регулирующих его отдачу. Вследствие этого температура их тела непостоянна, в известных пределах она следует за температурой окружающей среды. У карпа, линя, карася, находящихся в покое, температура тела соответствует температуре окружающей воды, а при плавании превышает ее на 0,2–0,3°С. Температура тела угря, у которого откладывается большое количество жира под кожей, может быть на 2,7°Свыше окружающей. У больных рыб температура тела повышается приблизительно на 2°С. Наибольшее превышение температуры тела над температурой воды 11°С зарегистрировано у тунца.
· Рыбы осваивают водоемы с самыми различными температурными режимами. В горячих источниках Калифорнии (t – 52°C) живет лукания. Угри были обнаружены при температуре 45°С; наряду с этим даллия обитает в промерзающих водоемах Крайнего Севера Азии (Чукотка) и Америки (Аляска). В нашей фауне исключительной холодостойкостью отличается карась – он способен, вмерзая зимой в лёд, оставаться живым (если только не промерзают полостные жидкости).
· Но это крайние примеры. Обычно в зависимости от пределов температуры, в которых происходит наиболее интенсивная нормальная жизнедеятельность, рыб разделяют на теплолюбивых и холодолюбивых.
· Теплолюбивые рыбы нашей фауны, такие, как сазан, карп, карась, линь, плотва, растительноядные, осетровые и др. , питаются наиболее интенсивно при температуре 17–28°С, при пониженной температуре пищевая активность ослабевает, а у ряда рыб на зиму прекращается, и они проводят зиму в малоподвижном состоянии в глубоких местах водоема; размножаются в теплое время года – весной и летом. Таким образом, обмен веществ у них наиболее эффективен при относительно высокой температуре (в определённых пределах).
· Для холодолюбивых рыб оптимальные температуры относительно низкие – 8–16°С; зимой они продолжают питаться; нерест происходит осенью и зимой (большинство лососевых – сиги, белорыбица, лосось, ручьевая форель и др.). При высоких температурах (более20°С) их активность падает, т. е. у этих рыб наиболее интенсивный обмен приспособлен к относительно низким температурам.
· Вся жизнедеятельность рыб (питание, рост, размножение и т. д. ) непосредственно определяется температурой воды, поэтому рыбы очень чувствительны к ее изменению.
· Общеизвестны скопления прудовых теплолюбивых рыб на наиболее прогреваемых участках водоемов и отход их на глубину во время похолодания; сардина концентрируется в слоях воды с температурой 18–26°С, а сайра – при 14–18°С. При организации промышленного лова обязательно учитывают распределение рыб в водоеме в соответствии с излюбленной ими температурой.
· Молодь рыб оказывается более устойчивой к колебаниям температуры, чем взрослые. Например, у карпа менее уязвимой к действию пониженной температуры является молодь сразу после выклева, т. е. когда она практически не может сменить биотоп и преимущественно висит, приклеившись к субстрату. В это время она может выносить даже заморозки до –0,1°С. Несколькими днями позже холодостойкость теряется, но молодь уже может перейти в более глубокое место.
· По способности переносить колебания температуры рыб разделяют на эвритермных (могут жить в широком диапазоне температур) и стенотермных (диапазон возможных температур узок). Эвритермные рыбы приспособились к жизни в изменяющихся условиях и переносят относительно широкие колебания температуры (щука, окунь, сазан, карп, карась, линь и др.)
· К стенотермным относят рыб, эволюция которых проходила в более или менее стабильных условиях, – обитателей тропической и полярной зон, а также больших глубин, где температура меняется мало.
· Однако и в пределах требуемой температуры повышение или понижение ее вызывает соответствующие сдвиги жизнедеятельности рыб. При этом изменяется реакция организма на одни и те же факторы среды: при повышении температуры увеличивается потребление кислорода, ускоряется развитие, усиливаются поиск, потребление и переваривание пищи – увеличивается желудочная секреция и моторная деятельность кишечника, ускоряется всасывание растворенных веществ из окружающей среды, повышается чувствительность к токсикантам и т. д. При этом большую роль играет состояние организма (возраст, степень половой зрелости и др. ). Чрезмерное охлаждение ведет к простуде (помутнение покровов, отслоение эпидермиса). Резкая смена температуры воды сказывается неблагоприятно на организме рыбы, поэтому необходимо выравнивать температуру воды при пересадках рыбы.
· Температура, выходящая из границ, допустимых для данного вида, вызывает шок и последующую гибель рыбы.
· Температура, при которой жизнь рыбы становится невозможной, называется пороговой. Приспособление организма к температурным условиям среды происходит на клеточном уровне. Пороговые температуры являются как бы пределом сопротивления клеток организма повреждающему действию внешней температуры. Поскольку эта способность у разных видов различна, то температурные пороги их неодинаковы. Однако на личинках карпа было показано, что пороговая температура может несколько меняться в зависимости от температуры выращивания. Более того, температурный режим содержания производителей сказывается на теплоустойчивости потомства. В рыбоводстве температура, при которой выращивают рыб, определяет быстроту развития, интенсивность питания и дыхания рыб, затраты кормов на прирост, направленность пластического обмена, скорость созревания, устойчивость потомства к колебаниям температуры и т. д.
· На стимулирующем воздействии повышенной температуры, если она не превышает максимальную, основываются и большие перспективы использования для рыбоводства теплых вод. Это касается прежде всего промышленных и сбросных вод электростанций и геотермальных вод.
· Большая растворяющая способность воды обеспечивает наличие различных как жизненно важных, так и вредных химических соединений во внешней среде и в организме рыбы.
· Растворенные в воде газы. Растворимость разных газов в воде не одинакова. Быстрее других растворяется двуокись углерода, далее – кислород, медленнее всех – азот. Поэтому для водоемов характерно иное соотношение газов, чем для атмосферы: в воде больше СО2 (4% против 0,05%) и кислорода (34% против 21%), но меньше азота (62% против 79%). При нормальных условиях отношение кислорода и азота в воде составляет почти 1: 2, в воздухе – 1: 4. Однако абсолютное содержание кислорода в воде в 20–30 раз меньше, чем в воздушной среде. Кислород из атмосферы диффундирует в воду медленно, поэтому содержание его убывает от поверхности к глубине. Многообразные физические и биологические процессы в водоеме (циркуляционные токи, ветровое перемешивание, жизнедеятельность растений и животных и т. д. ) являются причиной крайнего непостоянства кислородного режима малых водоемов.
· Подавляющее большинства рыб дышит растворенным в воде кислородом, поэтому содержание его в окружающей среде имеет для них первостепенное значение.
· Разные виды рыб нуждаются для нормального дыхания в разном количестве кислорода. Наиболее требовательным, обитателям холодных, проточных водоемов (например, лососевым) необходима концентрация 4,4–7,0 мг/л, окунь Perca fluviatilis и ёрш Acerina cernua могут жить при содержании кислорода 2,5 мг, а наиболее выносливые рыбы нашей фауны караси не испытывают угнетения при концентрации 0,3 мг/л.
· Зависимость жизнедеятельности рыб от содержания в воде кислорода особенно заметна в период эмбрионального развития. Концентрация кислорода очень сильно влияет на скорость развития и выживаемость эмбрионов: так, при содержании кислорода 1,2 мг/л развитие карпа длилось 120 ч, причем вылупилось 40% эмбрионов; при концентрации кислорода 9,0–12,0 мг/л длительность развития сократилась до 70–68 ч, а количество вылупившихся эмбрионов возросло до 92–98%.
· Морские рыбы более требовательны к содержанию кислорода в воде, чем пресноводные. В воде морей кислорода растворено много, а колебания его содержания малы. Непостоянство же кислородного режима малых водоемов, в которых нередки заморы – летние и зимние, – заставляет пресноводных рыб приспосабливаться к дефициту кислорода.
· Однако избыток кислорода в воде также неблагоприятен для рыб. При перенасыщении воды кислородом (>200%) у рыб появляются пузырьки газа в кровеносных сосудах, затем наступают судороги и смерть.
· Большое значение для нормальной жизнедеятельности рыб имеет содержание в воде двуокиси углерода. При повышенном содержании ее в воде падает способность крови поглощать из воды кислород, дыхание учащается, но газообмен становится менее интенсивным.
· Вода способна поглощать большое количество двуокиси угдерода: при t= 15°С в 1 л воды может раствориться больше 1 л СО2; однако в природе содержание свободной СО2 в воде ничтожно, так как она связывается кальцием. Если этого не происходит, то наступает отравление рыбы двуокисью углерода и замор. Способность противостоять повышению концентрации СО2 у разных видов рыб неодинакова. Так, форель более чувствительна, чем карп, карась или линь.
· Критическими уровнями СО2 в 1 л являются: для форели 120– 140мг, для толстолобика – 200 (молодь) – 300 (взрослая рыба), для карпа – 200, для линя – больше 400 мг.
· Значение рН, обусловленное концентрацией водородных ионов, является одним из важнейших абиотических факторов внешней среды, определяющим видовой состав и численность гидробионтов водоема. Воздействие рН на жизнедеятельность гидробионтов связано с тем, что способность гемоглобина использовать растворенный в воде кислород при различных концентрациях водородных ионов неодинакова. Вследствие этого изменение рН воды приводит к изменению интенсивности дыхания и кислородного порога.
· Наиболее благоприятно для дыхания большинства рыб значение рН, близкое к нейтральному. При сильных сдвигах рН в кислую и щелочную стороны (т. е. при увеличении или уменьшении концентрации водородных ионов) затрудняется дыхание, возрастает кислородный порог, ослабляется интенсивность питания.
· По отношению к колебаниям рН среды рыб делят на стено- и эвриионных. В воде морей рН изменяется мало (7,5–8,5), морские рыбы относятся к стеноионным. Пресные воды в отличие от морских характеризуются неустойчивостью рН. Это вызвано разнообразными факторами, направляющими ход биохимических процессов в водоеме: характером почв ложа и водосбора, химическим составом водоисточника, фотосинтетической деятельностью растений, особенно в период “цветения” воды, и т. д. В результате наблюдаются резкие годовые, сезонные и суточные колебания рН. Поэтому большинство пресноводных рыб приспособилось переносить значительные изменения рН и являются эвриионными.
· Однако возможные границы рН, в которых могут жить пресноводные рыбы, неодинаковы и при прочих равных условиях зависят прежде всего от вида. Из объектов рыборазведения наиболее выносливы карась и карп; щука переносит колебания рН в пределах 4,0–8,0, ручьевая форель – 4,5–9,5, карп – 4,3–10,8, карась выдерживает снижение рН до 4,5.
· Деятельность человека, изменяющая гидрохимический режим водоема, сильно отражается и на уровне рН. В водохранилищах, образованных при зарегулировании стока рек, концентрация водородных ионов колеблется в широких пределах. Сточные воды предприятий химической, металлургической, целлюлозной и другой промышленности содержат как кислоты, снижающие рН воды, так и щелочи, соду и другие компоненты, повышающие рН. Значение рН среды оказывает сильнейшее влияние на устойчивость рыб к различным токсическим веществам, входящим в состав промышленных стоков.
· Сероводород, образующийся в водоемах при отсутствии кислорода, оказывает на рыб губительное действие. Минимальная его летальная концентрация для рыб 1,0мг/л, однако разные рыбы реагируют неодинаково. Ручьевая форель при концентрации H2S 0,86 мг/л погибает через 24 ч, карп в то же время может жить при концентрации 6,3 мг/л.
· Солевой состав воды. Воздействие на рыб растворенных в воде солей заключается прежде всего в том, что от их количества зависит уровень осмотического давления.
· Большое значение имеет также и состав солей, так как они и непосредственно, и косвенно влияют на жизнедеятельность рыб.
· В организм рыб соли проникают через ротовую полость, жабры и кожу, причем проникновение солей через кожу зависит от плотности чешуйного покрова. Из общего количества поглощенных из окружающей воды солей фосфора у чешуйчатого карпа через жабры и ротовую полость проходит 93%, а через поверхность тела – 6,3%. У зеркального карпа на долю жабр и ротовой полости приходится 87,9%, а на поверхность тела– 12,1%. Попадая в организм, соли включаются в обмен веществ. Так, при увеличении концентрации солей фосфора в воде до 10 мг/л резко ускорялся рост молоди осетровых.
· Велико косвенное влияние солей на рыб. Количество и состав солей, выносимых с площади водосбора или внесенных при удобрении прудов, определяют богатство водоема биогенными веществами, а значит, создают основу для развития пищевых организмов для рыб (фитопланктон, зоопланктон, бентос), т. е. кормности водоема.
· Установлена прямая зависимость между наличием в воде фосфора, распределением планктона и уловами морских пелагических рыб.
· Воздействие человека на природу – развитие промышленности и судоходства, создание новых и реконструкция существующих водных систем, промышленные и бытовые стоки, интенсификация сельского хозяйства, включающая в широких масштабах применение химических средств защиты растений, удобрение полей и прудов, дезинфекция и мелиорация прудов и другие мероприятия – влечет за собой изменение режима естественных вод. Влияние этих факторов на гидробионтов может быть или прямым, или косвенным –через изменение температурного, газового, солевого режимов.
· В настоящее время загрязнение охватывает уже и океаны, но наиболее сильно оно проявляется во внутренних водоемах.
· Кислоты и щелочи сточных вод не только сдвигают рН до границ, смертельных для рыб, но и сами являются ядами, вызывая патоморфологические изменения органов (ожоги жабр и кожи, замедление роста) и гибель рыб.
· Летальными дозами являются содержание в 1 л воды 134 мг серной кислоты, 159 мг соляной, 200 мг азотной. Борная кислота в концентрации 62–500 мг/л снижала темп роста предличинок севрюги, а в концентрации 1500–2500 мг/л вызывала их гибель. Среди минеральных веществ стоков особенно ядовиты цианиды, соединения ртути, мышьяка, свинца, меди. Смертельными дозами для гольяна, карпа, горчака и дафний является содержание в 1 л KCN –0,06 мг, HgCl2 – 0,002 мг, NaHAsO3 – 0,5 мг. Соединения свинца вызывают гибель рыб в концентрации 10–150 мг/л, планктонных рачков – 0,5 мг/л. Гибель рыб могут вызвать соединения железа при содержании 0,2мг/л, алюминия 0,5мг/л, натрия 10–15 г/л, кальция 15 г/л и т. д.
· Удобрение площади водосбора и бесконтрольное внесение удобрений в пруды может создать в них избыток минеральных веществ. Наиболее опасна перегрузка водоема аммонийно-аммиачными удобрениями (селитра, нитрофоска, аммиачная вода и др.), так как аммиак и соли аммония являются ядами комбинированного действия: локального, нервно-паралитического и гемолитического. Поэтому, например, аммиак при даже относительно небольших концентрациях вызывает острое отравление рыб: голавля при 1,0–1,2мг NН3/л (при температуре 14°С и содержании кислорода 9–10мг/л), форели при 0,3–0,4 мг/л.
· Из органических веществ наиболее вредны синтетические моющие препараты, фенол, крезол и нафтеновые кислоты, обычно смертельные в концентрациях 10–100 мг/л.
· Воздействие токсикантов проявляется неодинаково и зависит от внешних и внутренних факторов. Большое значение имеет температура воды. При температуре 1°С летальная концентрация СО2для карпа 120 мг/л, при 30°С – 55–60 мг/л. Изменяется устойчивость организма рыб в связи с солевым составом воды. В мягкой воде токсичность кислот увеличивается, так как в ней очень быстро сдвигается рН. Различна сила влияния ядов в зависимости от возраста рыб, причем они наиболее чувствительны на ранних стадиях развития. Наиболее уязвимым становится организм в период, когда усилен обмен веществ, и, в частности, в период интенсивного питания рыб.
· Свет, звук, электропроводность, запах. Свет в воде поглощается довольно быстро, причем лучи разной части спектра проникают на разную глубину: красные и желтые лучи не проникают глубже 10м, синие и фиолетовые проходят глубже других – до 100 м.
· Освещенность резко убывает с глубиной и на больших глубинах (1,0–1,5км) царит полная темнота.
· Прозрачность пресноводных водоемов значительно меньше, чем морских, и колеблется сильнее: от прозрачных горных ручьев до мутных коричневых рек Азии или некоторых черных притоков Амазонки.
· К особенностям освещения приспособлены строение органов зрения рыб, степень развития других органов чувств, окраска покровов, наличие (или отсутствие) органов свечения и т. д. У видов, обитающих в поверхностных слоях, вырабатывается положительный фототаксис, а у придонных и сумеречных – отрицательный. С освещенностью связаны активность рыб, ход обменных процессов, половое созревание.
· Звук распространяется в воде в 4,5 раза быстрее, чем в воздухе, а поглощается в тысячу раз медленнее, т. е. слышен на десятки километров. Поэтому в воде его улавливать легче, чем в воздухе. По звукопроводимости ткани тела рыб близки к воде. В связи с этим даже относительно слабо развитый орган слуха рыб (внутреннее ухо)обеспечивает восприятие значительной информации из окружающей среды. Кроме того, в восприятии звуков принимают участие также органы боковой линии и плавательный пузырь (служащий резонатором).
· Электропроводность воды обусловлена тем, что большинство солей находится в ней в диссоциированном состоянии, в виде ионов. Многие рыбы воспринимают изменения электрического поля в воде, используют слабые электрические разряды для ориентировки, сигнализации, нападения.
· Искусственно создаваемое электрическое поле вызывает у рыб возбуждение, привлечение или электронаркоз (шок), в зависимости от расстояния до анода. Многие морские рыбы обнаруживают положительный электротаксис, пресноводные – отрицательный.
· Лов рыбы с помощью электротока применяют в малых водоемах (неспускных прудах). Соответствующие исследования ведутся в морском рыболовстве (с постоянным, переменным и импульсным токами).
· Запахи сохраняются в воде дольше и более стойко, чем в воздухе. Они являются источником разнообразной информации (в пищевом, нерестовом, оборонительном, стайном поведении и др. ). Рыбы воспринимают широкий спектр запахов, различают химические соединения различных классов (спирты, кетоны, эфиры, кислоты и др. ). Рыбы с хорошо развитой обонятельной чувствительностью (сом, налим, угорь, линь) сильно реагируют на запахи пищи, рыб своего и других видов и т. д.
· Благодаря восприятию видового запаха, свойственного слизи, рыбы отличают запах своей стаи (что позволяет им сохранить ее), обнаруживают они и запах рыб других видов, при этом мирные рыбы особенно чутко улавливают запах хищников – щуки, сома, ротана, а хищники – своих жертв: карасей, линей, карпов и особенно осетров. Акулы могут улавливать запах добычи на расстоянии до 500 м.
· Бич южноамериканских рек – пираньи; привлеченные запахом раненого животного, они молниеносно окружают его огромными стаями.
· Удивительная тонкость обоняния позволяет рыбам ориентироваться на запах родного водоема, который определяется метаболитами его обитателей.
· Определено, что вещества, выделяемые кожей морского льва в невообразимо слабой концентрации – 1: 80 000 000 000, вызывают испуг и тревогу рыб. Органы обоняния рыб обнаруживают в воде “вещество страха” и ферромоны.
· Высокая чувствительность обоняния рыб позволила использовать их в качестве живых индикаторов при очистке сточных вод предприятий нефтяной, химической и другой промышленности. В частности, эту роль выполняют золотые рыбки и зеркальный карп, живущие в отстойниках: если они становятся возбужденными, то это значит, что загрязнение воды усилилось. Способность рыб находить пищу по запаху используется в любительском рыболовстве (лов налима, бычка и других рыб на пахнущие приманки).
· Грунт и взвешенные в воде частицы. Большую роль в жизни рыб имеют взвешенные в воде частицы. Они определяют в значительной мере прозрачность воды и тем самым влияют на освещенность толщи воды и дна.
· Большая часть рыб в той или иной мере связана с дном водоема, т, е, с грунтом. Приспособления к жизни на дне развиваются в разных направлениях. Общеизвестна способность рыб менять окраску, внешние покровы донных рыб повторяют тон и рисунок дна, Закапывающиеся виды осваивают мягкие иловые участки, а живущие на каменистых грунтах обладают присосками и т. д.
· Среди пресноводных рыб при высыхании могут зарываться в ил водоема вьюн Misgurnus fossilis и карась Carassius carassius. Перекапывают ил в поисках пищевых организмов бентосоядные рыбы, многие лососевые перед нерестом разбрасывают хвостом гальку, устраивая “гнезда” (ямки) для икры.
· Защитными приспособлениями от избытка взвеси в воде у обитателей мутных вод являются уменьшение глаз (т. е. сокращение наиболее уязвимых участков поверхности тела) и усиленное выделение слизи, которая, осаждая муть, обеспечивает чистоту воды вокруг тела рыбы и таким образом улучшает условия дыхания.
Глава I. Строение и некоторые физиологические особенности рыб
· Строение и функции организма рыбы отражают его связь с водной средой.
· Специфические приспособления рыб весьма многообразны. Возникновение и характер их обусловлены чрезвычайным разнообразием среды и образа жизни.
Форма тела. Способы движения
· Форма тела должна обеспечивать рыбе возможность передвигаться в воде (среде значительно более плотной, чем воздух) с наименьшей затратой энергии и со скоростью, соответствующей ее жизненным потребностям.
· Форма тела, отвечающая этим требованиям, выработалась у рыб в результате эволюции: гладкое, без выступов, тело, покрытое слизью, облегчает движение; шеи нет; заостренная голова с прижатыми жаберными крышками и сжатыми челюстями рассекает воду; система плавников определяет движение в нужном направлении. В соответствии с образом жизни выделено до 12 различных типов формы тела. Наиболее характерные представлены на рис. 2.
· 1. Торпедовидный (веретенообразный). Тело рыб похоже на торпеду или веретено, оно хорошо обтекаемо, немного сжато с боков и утончается к хвосту. Рыбы приспособлены к быстрому длительному плаванию в толще воды. Это наилучшие пловцы, совершающие продолжительные миграции к местам нагула и к местам икрометания (нерестилищам): тунец, макрель, сельдь, треска, лососи.
· Такая же форма тела выработалась и у других хорошо плавающих водных животных, далеко отстоящих от рыб (китообразные и вымершие ихтиозавры).
· 2. Змеевидный. Тело вытянутое, змеевидное, округлое, на поперечном разрезе образует овал. Плавают, змеевидно изгибаясь всем телом. Это миноги, угри.
· 3. Лентовидный. Тело, подобное ленте, вытянуто вдоль, плоское с боков. Пловцы плохие, живут в спокойных водах больших глубин (сельдяной король, рыба-сабля).
· 4. Стреловидный. Тело удлиненное, сжато с боков, примерно одинаковой высоты; хвост сильный, голова заострена, спинной плавник сдвинут сильно назад. Эти рыбы продолжительных плаваний не совершают, но на небольшом расстоянии развивают огромную скорость, набрасываясь на добычу. Это хищники – щука, таймень.
· 5. Сплющенный. Здесь различают: а) симметрично-сжатую, лещевидную форму: тело высокое, сжатое с боков (лещ); б) несимметрично-сжатую: высокое, сжатое с боков тело несимметрично, глаза расположены на одной стороне (камбалы).
· Такая форма тела не способствует быстрому перемещению, эти рыбы плохие пловцы.
· 6. Плоский. Тело сплющено в дорсовентральном направлении (сверху вниз). Обычно двигаются мало, живут у дна (скаты).
· 7. Шаровидный. Тело в виде шара, иногда передний отдел окружен костным панцирем (кузовок). Самостоятельно передвигаться иногда почти не могут. Это скалозубы – Tetrodon, Diodon.
· Этими типами не исчерпывается многообразие форм тела рыб;у некоторых рыб форма тела является как бы промежуточной комбинацией нескольких типов.
· Внешние признаки рыб имеют большое значение при их определении. Основные части тела – голова, туловище, хвост, плавники – очень варьируют у разных видов по размерам, форме, соотношению.
· Форма головы очень разнообразна прежде всего в связи со строением ротового аппарата. Акулы – меч-рыба, пила-рыба, молот-рыба, игла, лопатонос своим названием обязаны видоизменениямчелюстей.
· Многие глубоководные рыбы имеют огромный (около 1/4 длины тела) рот, благодаря которому они могут захватывать добычу, большую, чем они сами.
· В прямой связи со способами питания находится положение рта (рис. 3). Различают рот верхний (планктоноядные рыбы), конечный (например, хищники), нижний (бентосоядные). Существуют и переходные формы – рот полуверхний, полунижний.
· Многие рыбы (осетровые, карповые) имеют выдвижной рот, благодаря которому они легко роются в иле, отыскивая пищу (рис. 4); у круглоротых рот превращен в присоску.
· У некоторых рыб (бычки, ротаны и др. ) голова вооружена шипами и колючками. Карп, сом и многие другие имеют усики (органы вкуса и осязания).
· Впереди глаз обычно расположены носовые, или обонятельные, отверстия (у круглоротых – непарные, у рыб – парные). У акул и скатов ноздри помещаются на нижней стороне головы, у остальных –на верхней.
· Позади глаз у акул, скатов и осетровых имеется небольшое отверстие – брызгальце (рудимент нефункционирующей жаберной щели). В задней части головы расположены жаберные отверстия, или щели. У круглоротых, скатов и акул количество этих отверстий соответствует количеству жаберных мешочков или жаберных дуг (5–7 с каждой стороны головы).
· У рыб жаберный аппарат усложнен: жаберные дуги (их 5) помещаются в жаберной полости, под прикрытием жаберной крышки. На внутренней вогнутой стороне четырех жаберных дуг имеются тычинки, образующие цедильный аппарат, на внешней выпуклой стороне – жаберные лепестки (органы дыхания).
· Число и форма жаберных тычинок сильно варьируют в зависимости от образа жизни и, в частности, от характера питания рыбы. Задняя часть жаберной крышки не прикреплена к голове, и поэтому по бокам головы образуются жаберные щели.
· Туловищный и хвостовой отделы тела рыб снабжены плавниками, благодаря которым рыба способна двигаться и удерживать равновесие. Лишенная плавников, она переворачивается брюшком вверх, поскольку центр тяжести помещается в спинной части.
· Плавники – это кожистые выросты, опирающиеся на костные плавниковые лучи. Различают парные плавники, соответствующие конечностям высших позвоночных, и непарные (вертикальные).
· Парные плавники – это грудные (p. pectoralis – Р) и брюшные (p. ventralis – V). Грудные плавники наиболее развиты у летающихрыб (позволяют им пролетать до 800 м) или ползающих по суше тропических Periophthalmys. Брюшные плавники сильно варьируют по положению: они могут смещаться вниз и вперед, под грудные (окунь) или даже на глотку (тресковые), превращаться в присоску (бычки, пинагор).
· Грудные и брюшные плавники выполняют функцию стабилизаторов, несущих плоскостей, рулей, иногда органов движения. С их помощью рыба поддерживает тело в нужном положении: при удалении их рыба плавает наклонно (в сторону удаленных плавников), а при перерезке грудных – хвостом вверх.
· Непарные плавники представлены спинным p. dorsalis – D, анальным p. analis – А и хвостовым p. caudalis – С.
· Количество спинных плавников варьирует (у карпа – 1, у судака –2, у трески – 3). У скумбриевых за двумя спинными и анальным плавниками находятся многочисленные маленькие добавочные плавнички. У некоторых рыб может быть не один, а два анальных плавника. Так же варьируют функции плавников. У рыбы-прилипало спинной плавник превращен в присоску, а у рыбы-парусника, достигая высоты 1,5 м и возвышаясь над водой, служит парусом. Непарные плавники обеспечивают устойчивость тела.
· Хвостовой плавник у подавляющего большинства рыб выполняет роль движителя, помогающего им плавать. Некоторые рыбы этого плавника не имеют (скаты).
· Форма хвостового плавника также связана с образом жизни рыбы.
· Неравнолопастный (гетероцеркальный) хвост большей частью сочетается с нижним ртом (акулы, осетровые).
· У летающих рыб сильнее развивается нижняя часть хвоста, и это помогает им выпрыгивать из воды. (У некоторых глубоководных рыб нижняя лопасть хвостового плавника больше длины тела. )
· Форма хвостового плавника связывается и со скоростью плавания: у наилучших пловцов – тунцов – хвост полулунный; медленнее плавает сельдь, имеющая вилообразный хвост; ещё медленнее –сазан, судак, лосось, обладающие выемчатым хвостом; у малоподвижного налима хвост округленный и т. д.
· У некоторых рыб (лососевые, корюшковые, хариусовые, косатковые, кошачьи сомы) позади спинного есть жировой плавник (adiposa), не имеющий костной опоры.
· В количестве и положении плавников возможны вариации. Парных плавников нет у круглоротых (миноги, миксины). Змеевидная форма тела обычно сопровождается потерей брюшных плавников (Anguillidae), а иногда и грудных (Muraenidae).
· При определении видовой принадлежности рыб особенно важен подсчет количества лучей в спинном (D) и анальном (А) плавниках, так как у костистых оно соответствует числу позвонков на определённом участке тела.
· Форма тела и его частей обусловливают возможность передвижения рыб (способы, скорости и т. д. ). Взаимодействие формы тела (обтекаемой, сплющенной или иной), размеров, формы и расположения плавников позволяет рыбе наилучшим образом использовать гидродинамические силы, возникающие при движении. У хороших пловцов (акулы, осетровые) подъемная сила увеличивается за счет выпуклой спинной и уплощенной брюшной поверхности тела, грудных плавников и рыла, направленных под углом к движению. Движения хвоста и хвостового плавника перемещают тело впереди изменяют направление движения в горизонтальной и вертикальной плоскостях.
· Различия в форме тела и образе жизни рыб привели к их различиям в способах движения. Основные из них таковы:
· 1. Движение при помощи боковых колебательных изгибов всего тела. Скорость движения невелика. Таким способом передвигаются придонные рыбы, имеющие удлиненное тело (угри, вьюны).
· 2. Движение при помощи частых боковых колебательных движений задней части тела. Передняя часть тела рассекает воду, задняя является движителем. Рыбы имеют компактное тело и мощный хвостовой стебель. К этой группе относится большинство рыб.
· Важная роль в обеспечении движения в водной толще принадлежит специальному гидростатическому органу – плавательному пузырю.
· У круглоротых и пластиножаберных (миноги, акулы) плавательного пузыря нет. Он появляется у более высокоорганизованных рыб.
· У костистых рыб, как правило, плавательный пузырь лежит в полости тела под позвоночником и почками над кишечником. Это однокамерный или двухкамерный орган, наполненный газами. Не имеют плавательного пузыря некоторые глубоководные рыбы, многие камбалы, а также рыбы, быстро меняющие глубину плавания (тунцы, скумбрии).
· В эмбриональном периоде у всех рыб плавательный пузырь соединен с кишечником тонкой трубкой, и первоначальное наполнение пузыря воздухом совершается через кишечник. Взрослых рыб по связи плавательного пузыря с кишечником разделяют на две группы. У открытопузырных трубка остается полой, так что связь пузыря с кишечником сохраняется в течение всей жизни и рыбы, заглатывая воздух, могут заполнять им пузырь (см. рис. 19). Это осетровые и низкоорганизованные костистые – лососевые, сельдевые, карповые. У закрытопузырных проток, соединяющий пузырь с кишечником, замыкается и пузырь оказывается изолированным от кишечника (все окуневые и пучкожаберные рыбы – колюшка, судак, окунь, морской конёк и др. ),
· Таким образом, после того как в первые дни жизни у личинок всех рыб плавательный пузырь наполняется воздухом в результате заглатывания его ртом, регуляция содержания газов в пузыре совершается у открытопузырных рыб путем периодических заглатываний воздуха, а у закрытопузырных – секрецией газов стенками пузыря. Газовая секреция у открытопузырных рыб развита слабо,
· Для выделения газов из крови в пузырь (газовая секреция) и наполнения его служат расположенные в передней части пузыря красные тела, или газовые железы, – скопления артериальных и венозных капилляров, называемых за определённый порядок расположения и большую концентрацию их на малой площади “чудесной сеточкой”. Например, у угря объём “чудесной сеточки” составляет 64 мм3 (объём одной капли воды); она состоит из 88 тыс. венозных и 116 тыс, артериальных капилляров, общая длина которых достигает 352–464 м.
· Удаление избытка газов из пузыря у открытопузырных рыб происходит через кровеносные сосуды стенки канала, соединяющего пузырь с кишечником, а также через рот. У закрытопузырных поглощение кислорода из пузыря совершается через овал – карманообразное, расположенное в задней части пузыря скопление капилляров. Изменение содержания газов в плавательном пузыре вызывает изменение плотности тела и связанной с ней плавучести.
· У личинок карпа первое наполнение плавательного пузыря воздухом происходит через 1–1,5 сут. после выклева. Лишенные этой возможности, они плохо растут и погибают на 10–14-й день.
· Кроме гидростатической плавательный пузырь выполняет ряд добавочных функций: барорецептора, добавочного органа дыхания, резонатора звуков, звукоиздающего органа.
Кожа. Чешуя. Органы свечения
· В кожном покрове рыб различают два слоя: наружный слой эпителиальных клеток, или эпидермис, и внутренний слой из соединительно тканных клеток – собственно кожа, дерма, кориум, кутис (рис. 5).
· Кожа подстилается рыхлой соединительнотканной прослойкой (подкожная соединительная ткань, подкожная клетчатка). У многих рыб в подкожной клетчатке откладывается жир.
· Эпидермис защищает организм от проникновения в него веществ извне. Он состоит из многослойного эпителия, форма клеток и количество слоев которого варьируют у разных рыб. Самый наружный слой эпителиальных клеток ороговевает, но в отличие от наземных позвоночных у рыб он не отмирает, сохраняя связь с живыми клетками. В течение жизни рыбы интенсивность ороговения эпидермиса не остается неизменной, наибольшей степени оно достигает у некоторых рыб перед нерестом: так, у самцов карповых и сиговых в некоторых местах тела (особенно на голове, жаберных крышках, боках и т. д. ) появляется так называемая жемчужная сыпь – масса мелких белых бугорков, придающих коже шероховатость. После нереста она исчезает.
· Кожа рыб отличается от кожи других позвоночных большим количеством слизи.
· Слизь образуется в специализированных железистых клетках, лежащих в эпидермисе. Это клетки трех форм: бокаловидные, колбовидные и зернистые. Присутствие всех форм определяет наибольшее количество слизи, при наличии одного вида клеток слизи выделяется значительно меньше.
· Интенсивность выделения слизи у разных рыб различна. Как правило, рыбы с хорошо развитым чешуйчатым покровом выделяют слизи меньше (лососевые, окуневые).
· Рыбы, лишенные чешуи или чешуя которых редуцирована (круглоротые, некоторые сомы, линь, вьюн), выделяют слизи очень много. С бактерицидными свойствами слизи линя связывают повышенную устойчивость его к заражению паразитами, тогда как другие представители семейства карповых такой устойчивостью не обладают.
· Роль слизи в жизненных отправлениях рыб очень велика: она уменьшает трение тела о воду (механическая защита), предотвращает попадание в организм паразитов и бактерий (бактерицидная защита), ускоряет свертывание крови в случаях ранений, способствует выведению веществ из организма, регулирует проникновение воды и солей (осмотическая регуляция), осаждает муть и выделяет специфический видовой запах.
· По некоторым данным, слизь некоторых рыб ядовита. Слизь миног вызывает у хищников нарушение пищеварения.
· Также в одноклеточных железах кожи образуются феромоны –летучие (пахучие) вещества, выделяемые в окружающую среду и воздействующие на рецепторы других рыб. Они специфичны для разных видов, даже близкородственных; в некоторых случаях определена их внутривидовая дифференцировка (возрастная, половая).
· У многих рыб, в том числе у карповых, образуется так называемое вещество страха (ихтиоптерин), которое выделяется в воду из тела пораненной особи и воспринимается ее сородичами как сигнал, извещающий об опасности.
· Светящиеся органы морских глубоководных рыб и ядовитые железы некоторых рыб, помещающиеся у основания шипов и плавниковых лучей (морской дракон), также являются железистыми образованиями эпидермиса.
· В нижних слоях эпидермиса в пограничных с ними слоях кориума залегают пигментные клетки– хроматофоры. Хроматофоры –звездчатые клетки, со множеством отростков, включающие зернышки пигмента. Они определяют все разнообразие окраски рыб, особенно яркой в тропиках. Различие в окраске достигается сочетанием разных хроматофоров: меланофоры имеют зерна черного пигмента, ксантофоры – жёлтого, эритрофоры – красного; гуанофоры или иридоциты не имеют пигментных зерен, но содержат кристаллики гуанина, благодаря которым рыба приобретает серебристую окраску.
· Интенсивность окраски определяется состоянием хроматофоров: при их расширении пигментные зерна растекаются на большее пространство и окраска тела становится яркой. Если хроматофоры сокращаются, пигментные зерна скапливаются в центре, оставляя большую часть клетки неокрашенной (рис. 6), и окраска тела бледнеет.
· Форма пигментных клеток постоянна и у близких видов сходна.
· Состояние пигментных клеток меняется в зависимости от внешних и внутренних факторов: температуры и газового режима водоема, возраста, пола, состояния организма (голод, размножение и т. п. ), эмоций (возбуждение, страх) и т. д.
· Окраска тела часто имеет приспособительное значение. У пелагических рыб обычно спина темная, а брюшко серебристо-белое, из-за чего рыба малозаметна и сверху и снизу.
· Общеизвестно приспособление окраски тела к цвету дна у камбал, бычков, морских игл: они могут повторять даже рисунок шахматной доски, положенной на дно. При этом основную роль играет нервная система, воспроизводящая зрительные раздражения и вызывающая соответствующую реакцию пигментных клеток (у ослепленных рыб кожа не меняла окраску).
· Изменение окраски в период размножения (появление брачного наряда), часто наблюдаемое в этот период различие в окраске самок и самцов происходят под влиянием гормональных факторов.
· Собственно кожа обеспечивает прочность покровов. Она состоит из нескольких слоев соединительной ткани (у костистых рыб, например, из трех), пронизанных нервами и капиллярами. В этом же слое залегают специализированные клетки – склеробласты, выделяющие секрет, который, застывая, образует чешую (кожный скелет), основным назначением которой является механическая защита тела.
· Все настоящие рыбы обладают чешуей, но степень развития её у разных рыб различна. Некоторые рыбы (например, сомы, скаты)в процессе эволюции утратили чешую, так что отсутствие ее у них является вторичным явлением.
· Размер чешуек сильно варьирует – от микроскопических у угрей до нескольких сантиметров у индийского усача. Также разнообразна форма чешуек.
· Различают три формы чешуи (рис. 7): плакоидная, ганоидная и костная. Плакоидная – наиболее древняя, сохранилась у хрящевых рыб (акулы, скаты). Состоит из пластинки, на которой возвышается шипик. Старые чешуйки сбрасываются, на их месте возникают новые. Ганоидная – преимущественно у ископаемых рыб. Чешуйки имеют ромбическую форму, тесно сочленяются одна с другой, так что тело оказывается заключенным в панцирь. Чешуйки со временем не меняются. Названием своим чешуя обязана ганоину (дентинообразному веществу), толстым слоем лежащему на костной пластинке. Среди современных рыб ее имеют панцирные щуки и многоперы. Кроме того, она имеется у осетровых в виде пластинок на верхней лопасти хвостового плавника (фулькры) и жучек, разбросанных по телу (модификация нескольких слившихся ганоидных чешуек). Постепенно видоизменяясь, чешуя теряла ганоин. У современных костистых рыб ганоина уже нет, чешуйки состоят из костных пластинок (костная чешуя). Эти чешуйки могут быть: циклоидными –округлыми, с гладкими краями (карповые) и ктеноидными – с зазубренным задним краем (окуневые). Обе формы родственны, но циклоидная как более примитивная встречается у низкоорганизованных рыб. Бывают случаи, когда в пределах одного вида самцы имеют ктеноидную, а самки – циклоидную чешую (камбалы рода Liopsetta), или даже у одной особи встречаются чешуйки обеих форм.
· Каждая такая чешуйка начинает образовываться в чешуйном кармашке – углублении в кориуме, затем свободный конец ее выходит из кармашка и накладывается на следующую чешуйку. Такое черепицеобразное расположение чешуек позволяет телу рыбы свободно изгибаться. Чешуя располагается рядами. Число рядов и количество чешуи в продольном ряду не изменяются с возрастом рыбы, поэтому они служат показателями при систематическом определении.
· В разрезе на каждой чешуйке различаются наружный покрышечный слой (преимущественно неорганического состава) и нижний базальный (имеющий характер кости). На верху покрышечного слоя концентрическими слоями располагаются ребрышки – склериты. Образование склеритов происходит периодически, поэтому их ряды подсчитывают при определении возраста рыб (годичные кольца) и обратного расчисления роста (см. с. 117).
· Кожа рыб быстро регенерирует. Через кожу происходит, с одной стороны, частичное выделение конечных продуктов обмена веществ, а с другой – поглощение некоторых веществ из внешней среды (кислород, угольная кислота, вода, сера, фосфор, кальций и другие элементы, играющие большую роль в жизнедеятельности). Большую роль играет кожа как рецепторная поверхность: в ней располагаются термо-, баро- хемо- и другие рецепторы. В толще кориума образуются покровные кости черепа и пояса грудных плавников.
СКЕЛЕТ
· Скелет у большей части рыб двоякий: наружный – защитный (чешуя) и внутренний – опорный.
· Внутренний скелет состоит из осевого скелета, скелета головы (черепной коробки, предохраняющей головной мозг и связанной с жаберным и челюстным аппаратом), скелета грудного (плечевого) и тазового поясов и плавников – парных и непарных (рис. 8).
· Осевой скелет может быть представлен хордой или позвоночником. У круглоротых, осетровых и двоякодышащих хорда сохраняется в течение всей жизни. У всех остальных рыб хорда имеется на ранних этапах развития, а у взрослых заменяется позвоночником, состоящим из позвонков.
· Черепная коробка соединена с позвоночником неподвижно. Шеи у рыб нет. Это вызывается спецификой образа жизни и среды обитания – необходимостью разрезать головой воду.
· В процессе эволюции происходило усложнение и окостенение скелета. У круглоротых хорда тянется от задней части черепа до хвоста в виде цельного несегментированного тяжа, состоящего из хрящевыхи соединительнотканных элементов (спинная струна), к которому сверху плотно прилегают хрящевые позвоночные дуги. Хорда осетровых также ещё не дифференцирована. У пластиножаберных (акуловых) рыб охрящевевшая оболочка хорды образует амфицельные (двояковогнутые) позвонки.
· Костистые рыбы имеют уже окостеневший позвоночник. В нём выделяют туловищный и хвостовой отделы. Туловищный отдел расчленен на типичные позвонки – амфицельные, в которых различают тело, верхнюю дугу с верхними (невральными) остистыми отростками (защищающими спинной мозг) и большие нижние дуги с нижними отростками. В туловищном отделе к позвоночнику (к поперечным отросткам или к телу позвонка) прикрепляются ребра. В хвостовом отделе поперечные отростки, смыкаясь, образуют нижнюю (гемальную) дугу, которая оканчивается нижним остистым отростком. В гемальном канале проходят хвостовые артерия и вена. Последний хвостовой позвонок уплощён и служит для прикрепления лучей хвостового плавника; он часто меняет обычную форму: удлиняется и загибается концом вверх, образуя уростиль.
· Количество позвонков определяется рядом внутренних и внешних факторов и служит систематическим признаком рыбы. Например, у северной сельди их 57, речного угря – 114, сома – 72, луны–рыбы – 17, судака – 44. В пределах вида известна зависимость количества позвонков (и лучей в грудном и анальном плавниках) от температуры: повышение температуры в период эмбриогенеза вызывает уменьшение их числа.
· Кроме ребер, опорную функцию у костистых рыб выполняют тонкие “мускульные” – межмышечные, или “туловищные”, косточки, пронизывающие мышцы. Эти косточки образованы окостеневшими сухожилиями. Больше всего их у карповых рыб.
· Череп рыб, так же как и осевой скелет, постепенно усложняется в процессе эволюции. У круглоротых нет черепной коробки; у них имеются отдельные, связанные соединительной тканью, хрящевые мозговые капсулы (обонятельная, слуховая, глазная).
· Хрящевые рыбы (акулы, скаты) имеют уже хрящевой череп, образованный сросшимися мозговыми капсулами, челюсти, вооруженные зубами. Челюстной аппарат соединяется с черепом. У осетровых, кроме того, возникает ряд кожных накладных костей, прикрывающих череп сверху, и настоящих костей, ограждающих череп снизу.
· Высшие костистые рыбы в эмбриональном периоде проходят все эти этапы: сначала у них закладывается и развивается хрящевой череп, позднее происходит окостенение его, причем наряду с замещающими костями возникают и покровные.
· В черепе костистых рыб выделяют два отдела: мозговой (осевой) и висцеральный (рис. 9). Многочисленные кости осевого отдела соединены неподвижно. Этим достигается надежная защита мозга. Висцеральный отдел черепа образуется челюстным и жаберным аппаратом. Он состоит из челюстной, подъязычной и пяти жаберных дуг, прикрытых жаберной крышкой.
· Жаберная крышка состоит из четырех костей (рис. 10): предкрышечной – ргаеoperculum, крышечной – operculum, межкрышечной – interoperculum и подкрышечной – suboperculum. Они прикрывают пять жаберных дуг. На внутренней стороне четырех дуг расположены жаберные тычинки, а на внешней – жаберные лепестки (органы дыхания). Кости челюстного и жаберного аппарата сочленованы с черепом подвижно и, будучи взаимосвязанными, двигаются согласованно при помощи мышц.
· На 5-й жаберной дуге нет лепестков, у некоторых рыб эта дуга превращается в нижнеглоточную кость, и на ней образуются глоточные зубы, которые достигают наибольшего развития у карповых. Глоточные зубы располагаются в 1–3 ряда. Форма и расположение глоточных зубов являются систематическими признаками.
· У карповых имеется также жерновок – роговое подушкообразное образование в верхней части глотки, служащее вместе с глоточными зубами для перетирания пищи.
· Грудной (плечевой) пояс состоит из трех основных костей: ключицы – cleitrum, лопатки – scapula и коракоида – coracoid. К нему прикрепляются грудные плавники. Он сочленен при помощи задневисочной кости с черепом.
· Тазовый пояс представлен у костистых рыб двумя срастающимися косточками, к которым прикрепляются лучи брюшных плавников. Он лежит в мышцах автономно, поэтому у некоторых видов может перемещаться далеко вперед, даже на горло, а иногда и вовсе исчезать. Скелет плавников являет собой опору, позволяющую рыбе пользоваться плавниками как рычагом или килем. У костистых рыб он представлен костными лучами, растягивающими кожистую плавательную перепонку.
· Различают лучи жесткие и мягкие (нечленистые и членистые), которые, в свою очередь, разделяются на разветвленные и неразветвленные. Лучи соединяются с поясами конечностей или при помощи специальных косточек (грудной плавник), или непосредственно (брюшной плавник). Количество лучей в плавниках D и А подсчитывают при определении рыб, так как у костистых оно соответствует числу позвонков на определённом участке тела,
· В соответствии с формой и строением различают несколько типов хвостового плавника (риc. 11).
· Все рыбы в эмбриональном и личиночном состоянии имеют прозрачную симметричную плавниковую кайму, огибающую конец позвоночника. Однако такая кайма у взрослых особей сохраняется только у круглоротых. У двоякодышащих позвоночник делит хвост на две равные части. Форма первично-симметричного хвоста (когда длинная ось тела делит его пополам) встречается редко.
· У древних групп рыб (акулы, осетровые) хвост несимметричный, неравнолопастный или гетероцеркальный: конец позвоночника резко загибается вверх, лопасти хвоста разные.
· У большинства костистых рыб (в том числе и у карпа) имеет место внутренняя асимметрия хвоста, образованная загнутым вверх уростилем, но при этом как вторичное явление развивается наружно-симметричная его форма. Такая форма называется ложно-симметричной.
· Осевой скелет и скелет поясов выполняют опорную функцию, кроме того, к ним прикрепляются двигательные мышцы.
Мышечная система и электрические органы
· Мышечную систему рыб, как и других позвоночных, разделяют мышечную систему тела (соматическую) и внутренних органов (висцеральную).
· В мышечной системе тела выделяют мускулы туловища, головы и плавников. Внутренние органы имеют свою мускулатуру.
· Мышечная система взаимосвязана со скелетом (опора при сокращении), с нервной системой (к каждому мышечному волокну подходит нервное волокно, и каждая мышца иннервируется определённым первом). Нервы, кровеносные и лимфатические сосуды располагаются в соединительнотканной прослойке мышцы. Сама соединительнотканная прослойка в мышцах рыб в отличие от мышц млекопитающих невелика.
· У рыб, как и других позвоночных, сильнее всего развита туловищная мускулатура. У настоящих рыб она представлена двумя большими тяжами, расположенными вдоль тела от головы до хвоста (большая боковая мышца – m. lateralis magnus) (рис. 12).
· Продольной соединительнотканной прослойкой эта мышца делится на спинную (верхнюю) и брюшную (нижнюю) части.
· Боковые мышцы разделены миотоммами (или миосептами) на миомеры, число которых соответствует количеству позвонков.
· Наиболее отчетливо миомеры видны у личинок рыб, пока их тела прозрачны.
· Мышцы правой и левой сторон, поочередно сокращаясь, сгибают хвостовой отдел тела и изменяют положение хвостового плавника, благодаря чему тело двигается вперед.
· Над большой боковой мышцей вдоль тела между плечевым поясом и хвостом у осетровых и костистых лежит прямая боковая поверхностная мышца (m. rectus lateralis, m. lateralis superficialis). У лососевых в ней откладывается большое количество жира. По нижней стороне тела тянется прямая брюшная мышца (m. rectus abdominalis); у некоторых рыб, например угрей, ее нет. Между ней и прямой боковой поверхностной мышцей располагаются косые мышцы (m. obliquus).
· Группы мышц головы управляют движениями челюстного и жаберного аппаратов (висцеральная мускулатура). Плавники имеют свою мускулатуру.
· Наибольшее скопление мускулов определяет и расположение центра тяжести тела; у большинства рыб он находится в спинной части. Деятельность туловищных мышц регулируется спинным мозгом и мозжечком, а висцеральная мускулатура иннервируется периферической нервной системой, возбуждаемой непроизвольно. Различают поперечно-полосатые и гладкие мышцы. К поперечно-полосатым относятся скелетные мышцы тела (туловищные) и мышцы сердца. Туловищные мышцы могут быстро и сильно сокращаться, однако скоро утомляются. Особенностью строения сердечных мышц является непараллельное расположение обособленных волокон, а разветвление их кончиков и переход из одного пучка в другой, что обусловливает непрерывную работу этого органа.
· Гладкие мышцы также состоят из волокон, но гораздо более коротких и не обнаруживающих поперечной исчерченности. Это мышцы внутренних органов и стенок кровеносных сосудов, имеющие периферическую (симпатическую) иннервацию.
· Поперечно-полосатые волокна, а следовательно, и мышцы делят на красные и белые, различающиеся, как следует из названия, цветом. Цвет обусловлен наличием миоглобина – белка, легко связывающего кислород; миоглобин обеспечивает дыхательное фосфорилирование, сопровождающееся выделением большого количества энергии.
· Красные и белые волокна различны по целому ряду морфофизиологических характеристик: цвету, форме, механическим и биохимическим свойствам (интенсивность дыхания, содержание гликогена и т. д. ).
· Волокна красной мышцы (m. lateralis superficialis) узкие, тонкие, интенсивно кровоснабжаемые, расположенные более поверхностно (вдоль тела от головы до хвоста), содержат в саркоплазме больше миоглобина; в них обнаружены скопления жира и гликогена. Возбудимость их меньше, отдельные сокращения длятся дольше, но протекают медленней; окислительный, фосфорный и углеводный обмен интенсивнее, чем в белых.
· В мышце сердца (красной) мало гликогена и много ферментов аэробного обмена (окислительный обмен). Она характеризуется умеренной скоростью сокращений и утомляется медленнее, чем белые мышцы.
· К широких, более толстых, светлых – белых волокнах m. lateralis magnins миоглобина мало; меньше в них гликогена и дыхательных ферментов. Углеводный обмен происходит преимущественно анаэробно, и количество выделяемой энергии меньше. Отдельные сокращения сравнительно быстры. Мышцы быстрее сокращаются и утомляются, чем красные. Лежат они более глубоко. Красные мышцы постоянно деятельны. Они обеспечивают длительную или непрерывную работу органов. Именно они поддерживают постоянное движение грудных плавников, обеспечивают изгибы тела при плавании и поворотах, непрерывную работу сердца.
· При быстром движении, бросках активны белые мышцы, при медленном – красные. Поэтому наличие красных или белых волокон (мышц) зависит от подвижности рыб: “спринтеры”-обладают почти исключительно белыми мышцами, у рыб, которым свойственны продолжительные миграции, кроме красных боковых мышц имеются добавочные красные волокна в белых мышцах.
· Основную массу мышечной ткани у рыб составляют белые мышцы. Например, у жереха, плотвы, чехони на их долю приходится 96.3, 95.2 и 94.9% соответственно.
· Белые и красные мышцы различаются по химическому составу. В красных мышцах содержится больше жира, тогда как в белых мышцах больше влаги и белка.
· Толщина (диаметр) мышечного волокна изменяется в зависимости от вида рыб, их возраста, величины, образа жизни; у прудовых рыб – от условий содержания.
· Например, у карпа, выращенного на естественной пище, диаметр мышечного волокна составляет (мкм): у мальков – 5–19, сеголетков 14–41, двухлетков – 25–50.
· Туловищная мускулатура образует основную массу мяса рыбы. Выход мяса в процентах от общей массы тела (мясистость) неодинаков у разных видов, а у особей одного вида различается в зависимости от пола, условий содержания и т. д.
· Мясо рыб усваивается быстрее, чем мясо теплокровных животных. Оно чаще бесцветно (судак) или имеет оттенки (оранжевый –у лососевых, желтоватый – у осетровых и т. д. ) в зависимости от наличия различных жиров.
· Основную массу белков мышц рыб составляют альбумины и глобулины (85%), всего же у разных рыб выделяют 4–7 фракций белков.
· Химический состав мяса (вода, жиры, белки, минеральные вещества) различен не только у разных видов, но и в разных частях тела. У рыб одного вида количество и химический состав мяса зависят от условий питания и физиологического состояния рыбы.
· В нерестовый период, особенно у проходных рыб, происходит расходование резервных веществ, наблюдается истощение и, как следствие, уменьшение количества жира и ухудшение качества мяса.
· У кеты, например, во время подхода к нерестилищам относительная масса костей увеличивается в 1,5 раза, кожи – в 2,5 раза. Мышцы оводняются – содержание сухого вещества снижается более чем в два раза; из мышц практически исчезают жир и азотистые вещества -рыба теряет до 98,4% жира и 57% белка.
· Особенность пищи и окружающей воды могут сильно изменить пищевую ценность рыбы: в заболоченных, тинистых или загрязненных нефтепродуктами водоемах рыбы имеют мясо с неприятным запахом.
· Качество мяса зависит от диаметра мышечного волокна, количества жира в мышцах и других факторов. В значительной мере оно определяется соотношением массы мышечной и соединительной ткани, по которому можно судить о содержании в мышцах полно ценных мышечных белков (по сравнению с неполноценными белками соединительнотканной прослойки).
· Это соотношение изменяется в связи с физиологическими особенностями организма и факторами внешней среды, в том числе с возрастом и условиями выращивания рыбы. В мышечных белках костистых рыб на белки саркоплазмы приходится 20–30%, белки миофибрилл – 60–70, белки стромы – около 2%.
· Все многообразие движений тела обеспечивает работа мышечной системы. Она главным образом обеспечивает и выделение тепла и электричества в организме рыбы.
· Своеобразно измененными мышцами являются электрические органы. Эти органы у большинства имеющих их рыб развиваются из зачатков поперечно-полосатой мускулатуры. Они расположены по бокам тела и у современных рыб состоят из множества мышечных пластинок (у электрического угря их около 6000), преобразованных в электрические пластинки, переслаиваемые студенистой соединительной тканью. Нижняя часть пластинки заряжена отрицательно, верхняя – положительно. Разряды происходят под действием импульсов продолговатого мозга.
· Электрические органы могут располагаться в разных частях тела, например у ската морской лисицы – на хвосте, у электрического сома – на боках.
· Генерируя электрический ток и воспринимая силовые линии, искаженные встречающимися на пути предметами, рыбы ориентируются в потоке, обнаруживают препятствия или добычу с расстояния в несколько метров даже в мутной воде. Органы зрения в этих условиях помочь им не могут и редуцируются.
· В соответствии со способностью к генерации электрических полей рыб разделяют на три группы:
· 1. Сильно электрические виды – имеют большие электрические органы, генерирующие разряды от 20 до 600 В. Основное назначение разрядов – нападение и оборона (электрический угорь, электрический скат, электрический сом).
· 2. Слабоэлектрические виды – имеют небольшие электрические органы, генерирующие разряды напряжением менее 17 В. Основное назначение разрядов – локация, сигнализация, ориентация (обитающие в мутных реках Африки многие мормириды, гимнотиды, некоторые скаты).
· 3. Неэлектрические виды – не имеют специализированных органов, но обладают электрической активностью. Генерируемые ими разряды распространяются на 10–15 м в морской воде и до 2 м в пресной. Основное назначение генерируемого электричества – локация, ориентация, сигнализация (многие морские и пресноводные рыбы: ставрида, атерина, окунь и др. ).
Пищеварительная система
· В пищеварительном тракте настоящих рыб различают ротовую полость, глотку, пищевод, желудок, кишечник (тонкая, толстая, прямая кишка, заканчивающаяся анусом). У акул, скатов и некоторых их рыб перед анусом имеется клоака – расширение, куда изливается прямая кишка и протоки мочевой и половой систем.
· В строении различных отделов имеется ряд особенностей. В ротовой полости рыб, как и других водных животных, нет слюнных желез, У высших позвоночных после смачивания пищи слюной в полости рта начинается ее частичная химическая обработка, у рыб ротовая полость служит или для отфильтровывания, отжатия пищи от воды (мирные), или для захватывания и удерживания добычи (хищники); железистые клетки ротовой полости и глотки выделяют слизь, которая не имеет пищеварительных ферментов, а способствует лишь проглатыванию пищи.
· Язык мощный, выдвижной имеют только круглоротые, у костистых рыб он не обладает собственной мускулатурой.
· В строении ротовой полости отражается приспособленность к определённому виду пищи.
· Рот и ротовая полость обычно снабжены зубами. У хищников они располагаются как на челюстях, так и на других костях полости рта, иногда даже на языке; они острые, часто крючкообразные, наклонены внутрь к глотке и служат для схватывания и удержания жертвы.
· Зубы рыб прирастают к костям или соединяются с ними подвижно. По мере снашивания они заменяются новыми. Наличием эмалевого колпачка и слоев дентина они напоминают зубы высших позвоночных.
· У мирных рыб (многие сельдевые, карповые и др.) на челюстях нет зубов.
· Тесно связаны со способом получения и видом пищи строение и подвижность челюстного и жаберного аппаратов. Жаберные щели, открываясь в глотку, связывают жаберную полость с пищеварительным трактом. Механизм питания координирован с дыхательным механизмом. Вода, всасываемая в рот при вдохе, несет и мелкие планктонные организмы, которые при выталкивании воды из жаберной полости (выдох) задерживаются в ней жаберными тычинками. Наиболее тонки, длинны и многочисленны (рис. 13) они у рыб, питающихся планктоном (планктонофагов), так что образуют фильтрующий аппарат (сельди, некоторые сиги); некоторые рыбы имеют для этой цели эпителиальные папиллы на жаберных дужках; у толстолобика, использующего фитопланктон, они даже срастаются в сеточку. Отцеженный таким образом комочек пищи направляется в пищевод.
· Хищные рыбы не нуждаются в отфильтровывании пищи, тычинки у них редкие, низкие, грубые, острые или крючковатые; они участвуют в удержании жертвы. У некоторых на жаберных дужках вместо тычинок имеются зубы. Но и у этих рыб захват и заглатывание добычи сообразуется с интенсивностью и ритмом дыхания.
· У некоторых бентосоядных рыб на задней жаберной дуге имеются глоточные зубы (рис. 14).
· Широкие, массивные, они служат для перетирания пищи. Наиболее сильно глоточные зубы развиты у карповых, камбаловых и некоторых других рыб. У карповых глоточные зубы разнообразны по строению. Кроме глоточных зубов в перетирании пищи у них участвует жерновик. Форма, количество и расположение глоточных зубов имеет систематическое значение.
· Следующий за глоткой пищевод, обычно короткий, широкий и прямой, с сильными мускулистыми стенками, проводит пищу в желудок. У открытопузырных рыб в пищевод открывается проток плавательного пузыря.
· Желудок чаще мешковидно расширен; наибольшей величины он достигает у хищников, что связано с размерами добычи. Однако не все рыбы имеют желудок. К безжелудочным относятся карповые, многие бычки и некоторые другие.
· В слизистой желудка имеются железистые клетки, специфические для этого отдела пищеварительного тракта, вырабатывающие соляную кислоту и пепсин, расщепляющий белок в кислой среде. Здесь у хищных рыб переваривается основная часть пищи.
· В начальную часть кишечника (тонкую кишку) впадают желчный проток и проток поджелудочной железы. По ним в кишечник попадают желчь и ферменты поджелудочной железы, под действием которых происходят расщепление белков до аминокислот, жиров – до глицерина и жирных кислот и расщепление полисахаридов до сахаров, главным образом глюкозы.
· В кишечнике, в условиях щелочной реакции, переваривание заканчивается. Особенно интенсивно оно происходит в переднем участке, имеющем больше пищеварительных соков. Здесь присутствует ряд ферментов, расщепляющих белки, жиры и углеводы. Важное значение имеет пристеночное пищеварение, которое, в частности, регулирует гидролиз крахмала.
· В кишечнике происходит всасывание питательных веществ, наиболее интенсивно – в заднем участке. Этому способствует складчатое строение его стенок, наличие в них ворсинкообразных выростов, пронизанных капиллярами и лимфатическими сосудами.
· У низших рыб (акулы, скаты, осетровые, двоякодышащие) в расширенном участке кишечника – толстой кишке – имеется спиральный клапан (образующий витки выроста стенки). Назначение его – увеличение внутренней (всасывающей) поверхности кишечника (рис. 15).
· У многих видов в начальной части кишечника помещаются слепые отростки – пилорические придатки, число которых сильно варьирует от 3 у окуня до 400 у лососей (у осетровых они срослись; рис. 16). Карповые, сомовые, щуки и ряд других рыб пилорических придатков не имеют. Пилорические придатки играют большую роль в пищеварении. Так, у радужной форели их общая длина более чем в 6 раз превышает длину кишечника, а их внутренняя поверхность в 3,2 раза превышает всасывающую поверхность переднего (тонкого) отдела кишечника. Гистологическое строение пилорических придатков одинаково со строением переднего отдела кишечника. Таким образом, при помощи пилорических придатков всасывающая поверхность кишечника увеличивается в несколько раз. В них происходит активный гидролиз белковых соединений; предполагают, что в пилорических придатках секретируются некоторые пищеварительные ферменты. У рыб, не имеющих желудка, кишечный тракт представляет собой большей частью недифференцированную трубку, суживающуюся к концу. У некоторых рыб, в частности у карпа, передняя часть кишечника расширена и напоминает по форме желудок. Однако это лишь внешняя аналогия – здесь нет характерных для желудка желез, вырабатывающих пепсин.
· У безжелудочных рыб переваривание пищи происходит в кишечнике, здесь же происходит всасывание питательных веществ.
· Строение, форма и длина пищеварительного тракта разнообразны в связи с характером пищи (объектами питания, их усвояемостью), особенностями переваривания. Наблюдается определённая зависимость длины пищеварительного тракта от рода пищи. Так, относительная длина кишечника (отношение длины кишечника к длине тела l)составляет у растительноядных пинагора и толстолобика 6–15, у всеядных карася и карпа 2-3, у хищных щуки, судака, окуня –1,2. Относительную длину кишечника рекомендуют использовать наряду с другими селекционными признаками в племенной работе с карпом.
· Печень крупная пищеварительная железа, по размерам уступающая у взрослых рыб только гонадам. Ее масса составляет уакул 14-25%, у костистых 1-8% массы тела. Это сложная трубчато-сетчатая железа, по происхождению связанная с кишечником (у зародышей является его слепым выростом).
· У большинства рыб, кроме некоторых лососевых, печень имеет многолопастную форму: в ней различают две, три, четыре a у карповых даже семь лопастей.
· В печеночной паренхиме тянутся печеночные артерии вены и желчные сосуды, собирающие желчь, вырабатываемую печеночными клетками.
· Желчные протоки проводят желчь в желчный пузырь (только у единичных видов его нет). Желчь благодаря щелочной реакции нейтрализует кислую реакцию желудочного сока. Она эмульгирует жиры, активирует липазу – фермент поджелудочной железы.
· Из пищеварительного тракта вся кровь медленно протекает через печень. В печеночных клетках кроме образования желчи происходит обезвреживание попавших с пищей чужеродных белков и ядов; откладывается гликоген, а у акул и тресковых (треска, налим и др. ) – жир и витамины. Пройдя через печень, кровь по печеночной вене направляется к сердцу.
· Объём печеночных клеток меняется под влиянием интенсивности синтеза и расхода углеводов, которые обусловлены, в свою очередь температурой окружающей среды, подвижностью, половой зрелостью рыб, интенсивностью питания и качеством пищи. Поэтому цвет и плотность ткани и общая масса печени сильно колеблются в зависимости от биологических особенностей рыбы и сезона года. При обильном полноценном питании печень приобретает красно-коричневый цвет с глянцевым отливом и некоторую упругость, масса ее увеличивается; у голодающих рыб она становится дряблой тусклой, мутной, жёлто-зелёной, ее объём и масса сильно уменьшаются. У прудовых карповых рыб к осени печень достигает максимальных размеров и массы и становится самой тяжелой из всех органов в полости тела; к весне, после длительного зимнего голодания, масса ее резко снижается. Сокращение объёма печеночных клеток после нереста установлено у радужной форели.
· Барьерная функция печени (очищение крови от вредных веществ путем образования безвредных соединений из поступающих ядов с находящимися в ней кислотами) обусловливает ее важнейшую рольне только в пищеварении, но и в кровообращении.
· Поджелудочная железа – сложная альвеолярная железа, также производная кишечника, является компактным органом только у акул и немногих других рыб. У большинства рыб она визуально не обнаруживается, так как диффузно внедрена в ткань печени (большей частью), и поэтому ее можно различить только на гистологических препаратах. В таком случае обе железы носят общее название hepatopancreas.
· У карповых рыб (линь, серебряный карась, сазан) поджелудочная железа представлена скоплением специализированных групп клеток, которые локализуются в печени, брыжейке и жировой ткани кишечника, а также в селезенке.
· В поджелудочной железе вырабатываются пищеварительные ферменты, действующие на белки, жиры и углеводы (трипсин, эрепсин, энтерокиназа, липаза, амилаза, мальтаза), которые выводятся в кишечник.
· У костистых рыб (впервые среди позвоночных) встречаются в паренхиме поджелудочной железы островки Лангерганса, в которых многочисленны клетки, синтезирующие инсулин, выделяемый прямо в кровь и регулирующий углеводный обмен.
· Таким образом, поджелудочная железа является железой внешней и внутренней секреции.
· Из мешкообразного впячивания спинной части начала кишечника образуется у рыб плавательный пузырь – орган, свойственный только рыбам.
Дыхательная система. Газообмен
· Эволюция рыб привела к появлению жаберного аппарата, увеличению дыхательной поверхности жабр, а отклонение от основной линии развития – к выработке приспособлений для использования кислорода воздуха. Большинство рыб дышит растворенным в воде кислородом, но есть виды, приспособившиеся частично и к воздушному дыханию (двоякодышащие, прыгун, змееголов и др. ).
· Основные органы дыхания. Основным органом извлечения кислорода из воды являются жабры.
· Форма жабр разнообразна в зависимости от видовой принадлежности и подвижности: это или мешочки со складочками (у рыбообразных), или пластинки, лепестки, пучки слизистой, имеющие богатую сеть капилляров. Все эти приспособления направлены на создание наибольшей поверхности при наименьшем объёме.
· У костистых рыб жаберный аппарат состоит из пяти жаберных дуг, располагающихся в жаберной полости и прикрытых жаберной крышкой. Четыре дуги на внешней выпуклой стороне имеют по два ряда жаберных лепестков, поддерживаемых опорными хрящами.
Таблица 1. Дыхательная поверхность жабр (по Строганову, 1962)
| . Виды рыб | Масса, г |
Дыхательная поверхность жабр
|
||
| Серебряный карась | 10,0 |
|
||
| Камбала | 135,0 |
|
||
| Окунь | 73,0 |
|
· Жаберные лепестки покрыты тонкими складками – лепесточками. В них и происходит газообмен. К основанию жаберных лепестков подходит приносящая жаберная артерия, ее капилляры пронизывают лепесточки; из них окисленная (артериальная)кровь по выносящей жаберной артерии попадает в корень аорты. Число лепесточков варьирует; на1 мм жаберного лепестка их приходится: у щуки – 15, камбалы – 28, окуня – 36. В результате полезная дыхательная поверхность жабр очень велика (табл. 1).
· Более активные рыбы имеют относительно большую поверхность жабр; у окуня она почти в 2,5 раза больше, чем у камбалы.
· Общая схема механизма дыхания у высших рыб представляется в следующем виде (рис. 17). При вдохе рот открывается, жаберные дуги отходят в стороны, жаберные крышки наружным давлением плотно прижимаются к голове и закрывают жаберные щели. Вследствие уменьшения давления вода всасывается в жаберную полость, омывая жаберные лепестки. При выдохе рот закрывается, жаберные дуги и жаберные крышки сближаются, давление в жаберной полости увеличивается, жаберные щели открываются и вода выжимается через них наружу. При плавании рыбы ток воды может создаваться за счет движения с открытым ртом.
· В капиллярах жаберных лепесточков из воды поглощается кислород (он связывается гемоглобином крови) и выделяются двуокись углерода, аммиак, мочевина. Большую роль играют жабры и в водно-солевом обмене, регулируя поглощение или выделение воды и солей. Замечательны приспособления для дыхания у рыб в эмбриональный период развития – у зародышей и личинок, когда жаберный аппарат ещё не сформирован, а кровеносная система уже функционирует. В это время органами дыхания служат: а) поверхность тела и система кровеносных сосудов Кювьеровы протоки, вены спинного и хвостового плавников, подкишечная вена, сеть капилляров на желточном мешке, голове, плавниковой кайме и жаберной крышке; б) наружные жабры (рис. 18). Это временные, специфические личиночные образования, исчезающие после образования дефинитивных органов дыхания. Чем хуже условия дыхания эмбрионов и личинок, тем сильнее развивается кровеносная система или наружные жабры. Поэтому у рыб, близких в систематическом отношении, но различающихся экологией нереста, степень развития личиночных органов дыхания различна.
· Дополнительные органы дыхания. К дополнительным приспособлениям, помогающим переносить неблагоприятные кислородные условия, относятся водное кожное дыхание, т. е. использование растворенного в воде кислорода при помощи кожи, и воздушное дыхание – использование воздуха при помощи плавательного пузыря, кишечника или через специальные добавочные органы (рис. 19).
· Дыхание через кожу тела – одна из характерных особенностей водных животных. И хотя у рыб чешуя затрудняет дыхание поверхностью тела, у многих видов роль так называемого кожного дыхания велика, особенно в неблагоприятных условиях. По интенсивности кожного дыхания пресноводных рыб делят на три группы:
· 1. Рыбы, приспособившиеся жить в условиях сильного дефицита кислорода. Это рыбы, населяющие хорошо прогреваемые, с повышенным содержанием органических веществ водоемы, в которых часто наблюдается недостаток кислорода. У этих рыб доля кожного дыхания в общем дыхании достигает 17–22%, у отдельных особей –42–80%. Это карп, карась, сом, угорь, вьюн. При этом рыбы, у которых кожа имеет наибольшее значение в дыхании, лишены чешуи или она мелкая и не образует сплошного покрова. Например, у вьюна 63%кислорода поглощается кожей, 37% – жабрами; при выключении жабр через кожу потребляется до 85% кислорода, а остальная часть поступает через кишечник.
· 2. Рыбы, испытывающие меньший недостаток кислорода и попадающие в неблагоприятные условия реже. К ним относятся обитающие у дна, но в проточной воде, осетровые – стерлядь, осетр, севрюга. Интенсивность кожного дыхания у них составляет 9–12% от общего.
· 3. Рыбы, не попадающие в условия значительного дефицита кислорода, живущие в проточных или непроточных, но чистых, богатых кислородом водах. Интенсивность кожного дыхания не превышает 3,3–9% от общего. Это сиги, корюшка, окунь, ёрш.
· Через кожу происходит также выделение углекислоты; так, у вьюна этим путем выделяется до 92% общего количества.
· При извлечении кислорода из воздуха во влажной атмосфере участвует не только поверхность тела, но и жабры. Важное значение при этом имеет температура.
· Наибольшей выживаемостью во влажной среде отличаются карась (11 сут.), линь (7 сут.), сазан (2 сут.), в то же время лещ, краснопёрка, уклея могут жить без воды всего несколько часов (при низкой температуре).
· Вьюн и угорь могут в течение нескольких дней жить вне воды при условии сохранения влажности кожи и жабр; это позволяет угрю переползать даже из одного водоема в другой.
· При перевозке живой рыбы без воды кожное дыхание почти целиком обеспечивает потребность организма в кислороде.
· У некоторых рыб, живущих в неблагоприятных условиях, выработались приспособления для дыхания кислородом воздуха. К ним прежде всего относится способ, специфичный для рыб, не свойственный другим позвоночным, – дыхание при помощи кишечника. В стенках кишечника образуются скопления капилляров. Воздух, заглатываемый ртом, проходит через кишечник, и в этих местах кровь поглощает кислород и выделяет двуокись углерода, при этом из воздуха поглощается до 50% кислорода. Такой вид дыхания свойствен вьюновым, некоторым сомовым и карповым рыбам; значение его у разных рыб неодинаково. Например, у вьюна в условиях большого недостатка кислорода именно этот способ дыхания становится почти равным жаберному.
· При заморах рыбы заглатывают ртом воздух; воздух аэрирует находящуюся в ротовой полости воду, которая проходит затем через жабры.
· Другим способом использования атмосферного воздуха служит образование специальных добавочных органов, например лабиринтового у лабиринтовых рыб, наджаберного у змееголова и др.
· Лабиринтовые рыбы имеют лабиринт – расширенный карманообразный участок жаберной полости, складчатые стенки которого пронизаны густой сетью капилляров, в которых происходит газообмен. Таким способом рыбы дышат кислородом атмосферы и могут находиться вне воды в течение нескольких дней (тропический окунь-ползун Anabas scandens выходит из воды и лазит по камням и деревьям; рис. 20).
· У змееголова выпячивание глотки образует наджаберную полость, слизистая оболочка ее стенок снабжена густой сетью капилляров. Благодаря наличию наджаберного органа он дышит воздухом и может находиться на мелководье при 30°С. Для нормальной жизнедеятельности змееголову, как и ползуну, нужен и растворенный в воде кислород, и атмосферный. Однако во время зимовки в прудах, покрытых льдом (зимовалах), он атмосферным воздухом не пользуется, а дышит только жабрами и кожей.
· Для использования кислорода воздуха служит рыбам и плавательный пузырь. Наибольшего развития как орган дыхания плавательный пузырь достигает у двоякодышащих рыб. Их ячеистый плавательный пузырь функционирует как легкое. При этом возникает “легочный круг” кровообращения.
· Состав газов в плавательном пузыре определяется как содержанием их в водоеме, так и состоянием рыбы.
· Подвижные и хищные рыбы имеют большой запас кислорода в плавательном пузыре, который расходуется организмом при бросках за добычей, когда поступление кислорода через органы дыхания оказывается недостаточным. В неблагоприятных кислородных условиях воздух плавательного пузыря у многих рыб используется для дыхания (в разной степени у разных видов).
· Карп и сазан, которые не имеют каких-либо специальных приспособлений для использования атмосферного воздуха, при нахождении вне воды частично поглощают кислород из плавательного пузыря.
· Осваивая различные водоемы, рыбы приспособились к жизни при разных газовых режимах. Наиболее требовательны к содержанию кислорода в воде лососевые, которым для нормальной жизнедеятельности нужна концентрация кислорода 4,4–7,0 мг/л; хариус, голавль, налим хорошо себя чувствуют при содержании в литре воды не менее 3,1 О2 мг/л, карповым обычно достаточно 1,9–2,5 мг/л. Каждому виду свойствен свой кислородный порог, т. е. минимальная концентрация кислорода, при которой рыба гибнет. Форель начинает задыхаться при содержании кислорода 1,9 мг/л, судак и лещ погибают при 1,2 мг/л, плотва и краснопёрка – при 0,25–0,3 мг/л; для выращенных на естественной пище карпов-сеголетков кислородный порог отмечен при 0,07–0,25 мг/л, а для двухлетков – 0,01 – 0,03 мг/л.
· Интенсивность дыхания определяется помимо видовой специфичности рядом биотических и абиотических факторов. Внутри одного вида она изменяется в зависимости от размера, возраста, подвижности, активности питания, пола, степени зрелости гонад, физико-химических факторов среды. По мере роста рыб активность окислительных, процессов в тканях уменьшается; созревание гонад, наоборот, вызывает увеличение потребления кислорода. Расход кислорода в организме самцов выше, чем у самок.
· На ритм дыхания, кроме концентрации в воде кислорода, влияют содержание СО2, рН, температура и т. д. Например, при температуре10°С и содержании кислорода 4,7 мг/л форель совершает 60–70 дыхательных движений в минуту, а при 1,2 мг/л частота дыхания возрастает до 140–160; карп при 10°С дышит почти вдвое медленнее, чем форель (частота дыхательных движений 30–40 раз в минуту), зимой он совершает в минуту 3–4 и даже 1–2 дыхательных движения. Как и резкий недостаток кислорода, на рыб губительно действует чрезмерное перенасыщение им воды.
· Отмечена гибель линей и карасей при насыщении воды кислородом до 150–200%: пузырьки газа покрывали жабры, были обнаружены под кожей, в органах, в артериях, кровь становилась пенистой; рыбы чувствовали себя плохо – дыхание их сначала учащалось, затем ослабевало, они выпрыгивали из воды и погибали в судорогах. Гибель годовиков карпа наблюдали после того, как в течение нескольких дней в пруду содержание кислорода достигало 200–240% насыщения. При этом зарегистрированы гиперемия и мраморный рисунок жабр, редкие дыхательные движения и медленные круговые или реже прямолинейные плавательные движения. Пересадка таких рыб в другой пруд не спасала их.
· Инкубация икры в перенасыщенной кислородом воде приводит к сильному увеличению отхода и количества уродов.
· Для нормального дыхания рыб очень важно содержание в воде СО2. При увеличении содержания свободной двуокиси углерода дыхание рыб становится невозможным, так как уменьшается способность гемоглобина крови связывать кислород, насыщение крови кислородом резко снижается и рыба задыхается. При высоком содержании СО2в атмосфере (1–5%) СО2 крови не может диффундировать наружу, а кровь не может принимать кислород даже из насыщенной кислородом воды.
Кровеносная система. Функции и свойства крови
· Главным отличием кровеносной системы рыб от других позвоночных является наличие одного круга кровообращения и двухкамерного сердца, наполненного венозной кровью (за исключением двоякодышащих и кистёперых).
· Сердце состоит из одного желудочка и одного предсердия и помещается в околосердечной сумке, сразу за головой, позади последних жаберных дуг, т. е. по сравнению с другими позвоночными сдвинуто вперед. Перед предсердием имеется венозная пазуха, или венозный синус, со спадающими стенками; через эту пазуху кровь поступает в предсердие, а из него – в желудочек.
· Расширенный начальный участок брюшной аорты у низших рыб (акулы, скаты, осетровые, двоякодышащие) образует сокращающийся артериальный конус, а у высших рыб – луковицу аорты, стенки которой сокращаться не могут. Обратному току крови препятствуют клапаны.
· Схема кровообращения в самом общем виде представлена следующим образом. Венозная кровь, заполняющая сердце, при сокращениях сильного мускульного желудочка через артериальную луковицу по брюшной аорте направляется вперед и поднимается в жабры по приносящим жаберным артериям. У костистых рыб их четыре с каждой стороны головы – по числу жаберных дуг. В жаберных лепестках кровь проходит через капилляры и, окисленная, обогащенная кислородом, направляется по выносящим сосудам (их также четыре пары) в корни спинной аорты, которые затем сливаются в спинную аорту, идущую вдоль тела назад, под позвоночником. Соединение корней аорты спереди образует характерный для костистых рыб головной круг. Вперед от корней аорты ответвляются сонные артерии.
· От спинной аорты идут артерии к внутренним органам и мускулатуре. В хвостовом отделе аорта переходит в хвостовую артерию. Во всех органах и тканях артерии распадаются на капилляры. Собирающие венозную кровь венозные капилляры впадают в вены, несущие кровь к сердцу. Хвостовая вена, начинающаяся в хвостовом отделе, войдя в полость тела, разделяется на воротные вены почек. В почках разветвления воротных вен образуют воротную систему, а выйдя из них, сливаются в парные задние кардинальные вены. В результате слияния вен задних кардинальных с передними кардинальными (яремными), собирающими кровь из головы, и подключичными, приносящими кровь из грудных плавников, образуется два Кювьерова протока, по которым кровь попадает в венозный синус. Кровь из пищеварительного тракта (желудка, кишечника) и селезенки, идущая по нескольким венам, собирается в воротную вену печени, разветвления которой в печени образуют воротную систему. Собирающая кровь из печени печеночная вена впадает прямо в венозный синус (рис. 21). В спинной аорте радужной форели обнаружена эластичная связка, выполняющая роль нагнетающего насоса, который автоматически увеличивает циркуляцию крови во время плавания, особенно в мускулатуре тела. Производительность этого “дополнительного сердца” зависит от частоты движений хвостового плавника.
· У двоякодышащих рыб появляется неполная перегородка предсердия. Это сопровождается и возникновением “лёгочного” круга кровообращения, проходящего через плавательный пузырь, превращенный в легкое.
· Сердце рыб относительно очень мало и слабо, гораздо меньше и слабее, чем у наземных позвоночных. Масса его обычно не превышает 0,33–2,5%, в среднем 1 % массы тела, тогда как у млекопитающих оно достигает 4,6%, а у птиц даже 10–16%.
· Кровяное давление (Па) у рыб низкое – 2133,1 (скат), 11198,8 (щука), 15998,4 (лосось), тогда как в сонной артерии лошади – 20664,6.
· Невелика и частота сокращений сердца – 18–30 ударов в минуту, причем она сильно зависит от температуры: при низких температурах у рыб, зимующих на ямах, она уменьшается до 1–2 ;у рыб, переносящих вмерзание в лед, пульсация сердца на этот период прекращается.
· Количество крови у рыб относительно меньше, чем у всех остальных позвоночных животных (1,1 – 7,3% от массы тела, в том числе у карпа 2,0–4,7%, сома – до 5, щуки – 2, кеты – 1,6, тогда как у млекопитающих – 6,8% в среднем). Это связано с горизонтальным положением тела (нет необходимости проталкивать кровь вверх) и меньшими энергетическими тратами в связи с жизнью в водной среде. Вода является гипогравитационной средой, т. е. сила земного притяжения здесь почти не сказывается.
· Морфологическая и биохимическая характеристика крови различна у разных видов в связи с систематическим положением, особенностями среды обитания и образа жизни. Внутри одного вида эти показатели колеблются в зависимости от сезона года, условий содержания, возраста, пола, состояния особей.
· Количество эритроцитов в крови рыб меньше, чем у высших позвоночных, а лейкоцитов, как правило, больше. Это связано, с одной стороны, с пониженным обменом рыб, а с другой – с необходимостью усилить защитные функции крови, так как окружающая среда изобилует болезнетворными организмами. По средним данным, в 1 мм3крови количество эритроцитов составляет (млн. ): у приматов –9,27; копытных– 11,36; китообразных – 5,43; птиц – 1,61–3,02; костистых рыб– 1,71 (пресноводные), 2,26 (морские), 1,49 (проходные).
· Количество эритроцитов у рыб колеблется в широких пределах, прежде всего в зависимости от подвижности рыб: у карпа – 0,84–1,89 млн. /мм3 крови, щуки – 2,08, пеламиды–4,12 млн. /мм3. Количество лейкоцитов составляет у карпа 20–80, у ерша – 178 тыс. /мм3. Клетки крови рыб отличаются большим разнообразием, чем у какой-либо другой группы позвоночных. У большинства видов рыб в крови имеются и зернистые (нейтрофилы, эозинофилы) и незернистые (лимфоциты, моноциты) формы лейкоцитов. Среди лейкоцитов преобладают лимфоциты, на долю которых приходится 80–95%, моноциты составляют 0,5–11%; среди зернистых форм преобладают нейтрофилы–13–31%; эозинофилы встречаются редко (у карповых, амурских растительноядных, некоторых окуневых).
· Соотношение разных форм лейкоцитов в крови карпа зависит от возраста и условий выращивания.
· Общее количество лейкоцитов в крови рыб сильно изменяется в течение года, у карпа оно повышается летом и понижается зимой при голодании в связи со снижением интенсивности обмена.
· Кровь окрашена гемоглобином в красный цвет, но есть рыбы и с бесцветной кровью. Так, у представителей семейства Chaenichthyidae (из подотряда нототениевых), обитающих в антарктических моряхв условиях низкой температуры (<2°С), в воде, богатой кислородом, эритроцитов и гемоглобина в крови нет. Дыхание у них происходит через кожу, в которой очень много капилляров (протяженность капилляров на 1 мм2 поверхности тела достигает 45 мм). Кроме того, у них ускорена циркуляция крови в жабрах.
· Количество гемоглобина в организме рыб значительно меньше, чему наземных позвоночных: на 1 кг массы тела у них приходится 0,5–4 г, тогда как у млекопитающих этот показатель возрастает до 5–25 г. У быстра передвигающихся рыб обеспеченность гемоглобином выше, чем у малоподвижных (у проходного осетра 4 г/кг, у налима 0,5 г/кг). Количество гемоглобина в крови рыб колеблется в зависимости от сезона (у карпа повышается зимой и понижается летом), гидрохимического режима водоема (в воде с кислым значением рН, равным 5,2, количество гемоглобина в крови возрастает), условий питания (карпы, выращенные на естественной пище и дополнительных кормах, имеют разную обеспеченность гемоглобином). Ускорение темпа роста рыб коррелирует с повышенной обеспеченностью их организма гемоглобином.
· Способность гемоглобина крови извлекать кислород из воды у разных рыб неодинакова. У быстро плавающих рыб – макрели, трески, форели – гемоглобина в крови много, и они очень требовательны к содержанию кислорода в окружающей воде. У многих морских придонных рыб, а также угря, карпа, карасей и некоторых других, наоборот, гемоглобина в крови мало, но он может связывать кислород из среды даже с незначительным количеством кислорода.
· Например, судаку для насыщения крови кислородом (при 16°С) необходимо содержание в воде 2,1–2,3 О2 мг/л; при наличии в воде 0,56–0,6 О2 мг/л кровь начинает его отдавать, дыхание оказывается невозможным и рыба гибнет. Лещу при этой же температуре для полного насыщения гемоглобина крови кислородом достаточно присутствия в литре воды 1,0–1,06 мг кислорода.
· Чувствительность рыб к изменениям температуры воды также связана со свойствами гемоглобина: при повышении температуры воды потребность организма в кислороде увеличивается, но способность гемоглобина связывать его падает.
· Угнетает способность гемоглобина связывать кислород и углекислота: для того чтобы насыщенность кислородом крови угря достигла 50% при содержании в воде 1% СО2, необходимо давление кислорода в 666,6 Па, а в отсутствии СО2 для этого достаточно давления кислорода почти вдвое меньшего – 266,6– 399,9 Па.
· Группы крови у рыб впервые были определены на байкальском омуле и хариусе в 30-х годах. К настоящему времени установлено, что групповая антигенная дифференцировка эритроцитов широко распространена; выявлено 14 систем групп крови, включающих более 40 эритроцитарных антигенов. При помощи иммуносерологических методов изучается изменчивость на разных уровнях; выявлены различия между видами и подвидами и даже между внутривидовыми группировками у лососевых (при изучении родства форелей), осетровых (при сравнении локальных стад) и других рыб.
· Кровь, будучи внутренней средой организма, содержит в плазме белки, углеводы (гликоген, глюкоза и др.) и другие вещества, играющие большую роль в энергетическом и пластическом обмене, в создании защитных свойств. Уровень этих веществ в крови зависит от биологических особенностей рыб и абиотических факторов, а подвижность состава крови позволяет использовать ее показатели для оценки физиологического состояния.
· Костного мозга, являющегося основным органом образования форменных элементов крови у высших позвоночных, и лимфатических желез (узлов) у рыб нет.
· Кроветворение у рыб по сравнению с высшими позвоночными отличается рядом особенностей:
· 1. Образование клеток крови происходит во многих органах. Очагами кроветворения у рыб являются: жаберный аппарат (эндотелий сосудов и ретикулярный синцитий, сосредоточенный у основания жаберных лепестков), кишечник (слизистая), сердце (эпителиальный слой и эндотелий сосудов), почки (ретикулярный синцитий между канальцами), селезёнка, сосудистая кровь, лимфоидный орган (скопления кроветворной ткани – ретикулярного синцития – под крышей черепа). На отпечатках этих органов видны кровяные клетки разных стадий развития.
· 2. У костистых рыб наиболее активно гемопоэз происходит в лимфоидных органах, почке и селезенке, причем главным органом кроветворения являются почки (передняя часть). В почках и селезенке происходит как образование эритроцитов, лейкоцитов, тромбоцитов, так и распад эритроцитов.
· 3. Наличие в периферической крови рыб и зрелых и молодых эритроцитов является нормальным и не служит патологическим показателем в отличие от крови взрослых млекопитающих.
· 4. В эритроцитах, как и у других водных животных, в отличие от млекопитающих имеется ядро.
· Селезёнка рыб располагается в передней части полости тела, между петлями кишечника, но независимо от него. Это плотное компактное тёмно-красное образование различной формы (шарообразной, лентовидной), но чаще вытянутой. Селезёнка быстро меняет объём под влиянием внешних условий и состояния рыбы. У карпа она увеличивается зимой, когда в связи с пониженным обменом веществ ток крови замедляется и она скапливается в селезенке, печени и почках, которые служат депо крови, тоже наблюдается при острых заболеваниях. При недостатке кислорода, перевозке и сортировке рыбы, облове прудов запасы крови из селезенки поступают в кровяное русло. Изменение размеров селезенки в связи с периодами усиленной активности установлено на ручьевой и радужной форелях и других рыбах.
· Одним из важнейших факторов внутренней среды является осмотическое давление крови, так как от него зависит в значительной степени взаимодействие крови и клеток тела, водный обмен в организме и т. д.
· Лимфатическая система рыб не имеет желез. Она представлена рядом парных и непарных лимфатических стволов, в которые лимфа собирается из органов и по ним же выводится в конечные участки вен, в частности в Кювьеровы протоки.
Выделительная система и осморегуляция
· В отличие от высших позвоночных, имеющих компактную тазовую почку (метанефрос), рыбы обладают более примитивной туловищной почкой (мезонефрос), а их зародыши – предпочкой (пронефрос). У некоторых видов (бычок, атерина, бельдюга, кефаль) предпочка в том или ином виде выполняет выделительную функцию и у взрослых особей; у большинства же взрослых рыб функционирующей почкой становится мезонефрос.
· Почки – парные, вытянутые вдоль полости тела темно-красные образования, плотно прилегающие к позвоночнику, над плавательным пузырем (рис. 22). В почке выделяют передний отдел (головная почка), средний и задний. Артериальная кровь поступает в почки по почечным артериям, венозная по воротным венам почек.
· Морфофизиологическим элементом почки является извитой почечный мочевой каналец, один конец которого расширяется в мальпигиево тельце, а другой отходит к мочеточнику. Железистые клетки стенок секретируют продукты азотистого распада (мочевину) , которые попадают в просвет канальцев. Здесь же, в стенках канальцев, происходит обратное всасывание воды, сахаров, витаминов из фильтрата мальпигиевых телец.
· Мальпигиево тельце – клубочек артериальных капилляров, охватываемый расширенными стенками канальца, – образует боуменову капсулу. У примитивных форм (акулы, скаты, осетровые) перед капсулой от канальца отходит мерцательная воронка. Мальпигиев клубочек служит аппаратом фильтрации жидких продуктов обмена. В фильтрат попадают как продукты обмена, так и важные для организма вещества. Стенки почечных канальцев пронизаны капиллярами воротных вен и сосудов из боуменовых капсул.
· Очищенная кровь возвращается в сосудистую систему почек (почечную вену), а отфильтрованные из крови продукты обмена и мочевина выводятся через каналец в мочеточник. Мочеточники изливаются в мочевой пузырь (мочевой синус), а затем моча выводится наружу у самцов большинства костистых рыб через мочеполовое отверстие позади ануса, а у самок костистых и самцов лососевых, сельдей, щуки некоторых других – через анальное отверстие. У акул и скатов мочеточник открывается в клоаку.
· В процессах выделения и водно-солевого обмена кроме почек принимают участие кожа, жаберный эпителий, пищеварительная система (см. ниже).
· Жизненная среда рыб – морские и пресные воды – всегда имеет большее или меньшее количество солей, поэтому осморегуляция является важнейшим условием жизнедеятельности рыб.
· Осмотическое давление водных животных создается давлением их полостных жидкостей, давлением крови и соков тела. Определяющая роль в этом процессе принадлежит водно-солевому обмену.
· Каждая клетка тела имеет оболочку: она полупроницаема, т. е. по-разному проницаема для воды и солей (пропускает воду и солеизбирательно). Водно-солевой обмен клеток определяется в первую очередь осмотическим давлением крови и клеток.
· По уровню осмотического давления внутренней среды по отношению к окружающей воде рыбы образуют несколько групп:
· у миксин полостные жидкости изотоничны окружающей среде;
· у акул и скатов концентрация солей в жидкостях тела и осмотическое давление немного выше, чем в морской воде, или почти равно ему (достигается за счет разницы солевого состава крови и морской воды и за счет мочевины);
· у костистых рыб – и морских и пресноводных (как и у более высоко организованных позвоночных) – осмотическое давление внутри тела не равно осмотическому давлению окружающей воды. У пресноводных рыб оно выше, у морских рыб (как и у других позвоночных) ниже, чем в окружающей среде (табл. 2).
Таблица 2 Величина депрессии крови для крупных групп рыб (по Строганову, 1962) ..
| Группа рыб | Депрессия Д° Кровь | Депрессия Д° Внешняя среда | Среднее осмотическое давление, Па Кровь | Среднее осмотическое давление, Па Внешняя среда |
| Костистые: морские | 0,73 | 1,90-2,30 |
8,9 • 105 |
25,1 • 105 |
| Костистые: пресноводные | 0,52 | 0,02-0,03 |
6,4 • 105 |
0,3 • 105 |
· Если в организме поддерживается определённый уровень осмотического давления жидкостей тела, то условия жизнедеятельности клеток становятся более стабильными и организм меньше зависит от колебаний внешней среды. Настоящие рыбы обладают этим свойством – сохранять относительное постоянство осмотического давления крови и лимфы, т. е. внутренней среды; поэтому они относятся к гомойосмотическим организмам (от греч. “гомойос” – однородный) .
· Но у разных групп рыб эта независимость осмотического давления выражается и достигается по-разному,
· У морских костистых рыб общее количество солей в крови значительно ниже, чем в морской воде, давление внутренней среды меньше давления внешней, т. е. их кровь гипотонична по отношению к морской воде. Ниже приведены величины депрессии крови рыб (по Строганову, 1962):
| Вид рыбы | Депрессия среды Д° |
|
Морские: треска балтийская камбала морская скумбрия форель радужная налим |
0,77 0,70 0,73 0,52 0,48 |
|
Пресноводные: карп линь щука |
0,42 0,49 0,52 |
|
Проходные: угорь в море “ в реке севрюга в море “ в реке |
0,82 0,63 0,64 0,44 |
· У пресноводных рыб количество солей в крови выше, чем в пресной воде. Давление внутренней среды больше давления внешней, их кровь гипертонична.
· Поддержание солевого состава крови и давления ее на нужном уровне обусловливается деятельностью почек, особых клеток стенок почечных канальцев (выделение мочевины), жаберных лепестков (диффузия аммиака, выделение хлоридов), кожных покровов, кишечника, печени.
· У морских и пресноводных рыб осморегуляция совершается разными способами (специфическая деятельность почек, различная проницаемость покровов для мочевины, солей и воды, различная деятельность жабр в морской и пресной воде).
· У пресноводных рыб (с гипертонической кровью), находящихся в гипотонической среде, разница осмотического давления внутри и вне организма приводит к тому, что вода извне непрерывно поступает внутрь организма – через жабры, кожу и ротовую полость (рис. 23).
· Во избежание чрезмерного обводнения, для сохранения водно-солевого состава и уровня осмотического давления возникает необходимость вывода из организма лишней воды и одновременного удержания солей. В связи с этим у пресноводных рыб мощное развитие получают почки. Количество мальпигиевых клубочков и почечных канальцев у них велико; мочи они выделяют гораздо больше, чем близкие морские виды. Данные о количестве мочи, выделяемой рыбами в сутки, представлены ниже (по Строганову, 1962):
| Вид рыбы | Количество мочи, мл/кг массы тела |
|
Пресноводные: карп форель сом карликовый |
50–120 60– 106 154 – 326 |
|
Морские: бычок морской черт |
3–23 18 |
|
Проходные: угорь в пресной воде “ в море |
60–150 2–4 |
· Утрата солей с мочой, экскрементами и через кожу восполняется у пресноводных рыб за счет получения их с пищей благодаря специализированной деятельности жабр (жабры поглощают из пресной воды ионы Na и Сl) и поглощением солей в почечных канальцах.
· Морские костистые рыбы (с гипотонической кровью), находящиеся в гипертонической среде, постоянно теряют воду – через кожу, жабры, с мочой, экскрементами. Предотвращение обезвоживания организма и сохранение осмотического давления на нужном уровне (т. е. ниже, чем в морской воде) достигаются тем, что они пьют морскую воду, которая всасывается через стенки желудка и кишечника, а избыток солей выделяется кишечником и жабрами.
· Угорь и морской бычок-подкаменщик в морской воде ежедневно пьют 50–200 см3 воды на 1 кг массы тела. В условиях опыта при пре прекращении подачи воды через рот (закрытый пробкой) рыба теряла 12%– 14% массы и на 3–4-й день погибала.
· Морские рыбы выделяют очень мало мочи: в почках у них немного мальпигиевых клубочков, у некоторых их нет совсем и есть только почечные канальцы. У них уменьшена проницаемость кожи для солей, жабры выделяют наружу ионы Na и Сl. Железистые клетки стенок канальцев увеличивают выделение мочевины и других продуктов азотистого обмена.
· Таким образом, у непроходных рыб – только морских или только пресноводных – действует какой-нибудь один, специфический для них способ осморегуляции.
· Эвригалинные организмы (т. е. выдерживающие значительное колебание солености), в частности проходные рыбы, проводят часть жизни в море, а часть – в пресной воде. При переходе из одной среды в другую, например во время нерестовых миграций, они переносят большие колебания солености. Это возможно благодаря тому, что проходные рыбы могут переходить с одного способа осморегуляции на другой. В морской воде у них действует такая же система осморегуляции, как у морских рыб, в пресной – как у пресноводных, так что их кровь в морской воде гипотонична, а в пресной – гипертонична.
· Их почки, кожа и жабры могут функционировать двояко: почки имеют почечные клубочки с почечными канальцами, как у пресноводных рыб, и только почечные канальцы, как у морских. Жабры снабжены специализированными клетками (так называемые клетки Кейс-Вильмера), способными поглощать и выделять Сl и Na (тогда как у морских или пресноводных рыб они действуют только в одном направлении). Изменяется и количество таких клеток. При переходе из пресной воды в море в жабрах японского угря возрастает количество клеток, выделяющих хлориды. У речной миноги при подъеме из моря в реки количество мочи, выделяемой в течение суток, увеличивается до 45% по сравнению с массой тела.
· У некоторых проходных рыб большую роль в регуляции осмотического давления играет слизь, выделяемая кожей.
· Передний отдел почки – головная почка – выполняет не выделительную, а кроветворную функцию: в него не заходит воротная вена почек, а в составляющей ее лимфоидной ткани образуются красные и белые кровяные клетки и разрушаются отжившие эритроциты.
· Как и селезёнка, почки чутко отражают состояние рыбы, уменьшаясь в объёме при недостатке кислорода в воде и увеличиваясь при замедлении обмена (у карпа – во время зимовки, когда ослабляется деятельность кровеносной системы), в случае острых заболеваний и т. д.
· Очень своеобразна дополнительная функция почек у колюшки, строящей для нереста гнездо из кусочков растений: перед нерестом почки увеличиваются, в стенках почечных канальцев вырабатывается большое количество слизи, которая в воде быстро затвердевает и скрепляет гнездо.
Половая система
· Развитие мочеполовой системы в эволюции рыб привело к обособлению половых протоков от выделительных.
· У круглоротых специальных половых протоков нет. Из разрывающейся половой железы половые продукты выпадают в полость тела, из нее – через половые поры – в мочеполовой синус, а затем через мочеполовое отверстие выводятся наружу.
· У хрящевых рыб половая система связана с выделительной. У самок большинства видов яйца выводятся из яичников по мюллеровым каналам, выполняющим роль яйцеводов и открывающимся в клоаку; вольфов канал является мочеточником. У самцов вольфов канал служит семяпроводом и через мочеполовой сосочек также открывается в клоаку.
· У костистых рыб вольфовы каналы служат мочеточниками, мюллеровы каналы у большинства видов редуцируются, половые продукты выводятся наружу через самостоятельные половые протоки, открывающиеся в мочеполовое или половое отверстие.
· У самок (большинства видов) зрелые яйца выводятся из яичника наружу через короткий проток, образованный оболочкой яичника. У самцов канальцы семенника соединяются с семяпроводом (не связанным с почкой), который открывается наружу мочеполовым или половым отверстием.
· Половые железы, гонады – семенники у самцов и яичники или ястыки у самок – лентовидные или мешковидные образования, висящие на складках брюшины – брыжейке – в полости тела, над кишечником, под плавательным пузырем. Строение гонад, сходное в основе, у разных групп рыб имеет некоторые особенности, У круглоротых половая железа непарная, у настоящих рыб гонады большей частью парные. Вариации в форме гонад у различных видов главным образом выражаются в частичном или полном слиянии парных желез в одну непарную (самки трески, окуня, бельдюги, самцы песчанки)или в ясно выраженной асимметрии развития: часто гонады бывают разные по объёму и массе (мойва, серебряный карась и др. ), вплоть до полного исчезновения одной из них. С внутренней стороны стенок яичника в щелевидную полость его отходят поперечные яйце несущие пластинки, на которых развиваются половые клетки. Основу пластинок составляют соединительнотканные тяжи с многочисленными ответвлениями. Вдоль тяжей проходят сильно разветвленные кровеносные сосуды. Зрелые половые клетки выпадают с яйценесущих пластинок в полость яичника, которая может быть расположена в центре его (например, окуневые) или сбоку (например, карповые).
· Яичник непосредственно сливается с яйцеводом, выводящим яйца наружу. У некоторых форм (лососевые, корюшковые, угревые) яичники не замкнуты и зрелые яйца выпадают в полость тела, а уже из неё через специальные протоки выводятся из организма. Семенники большинства рыб – парные мешковидные образования. Зрелые половые клетки по выводным протокам – семяпроводам – выводятся во внешнюю среду через специальное половое отверстие (у самцов лососей, сельдей, щуки и некоторых других) или через мочеполовое отверстие, расположенное позади ануса (у самцов большинства костистых рыб).
· У акул, скатов, химер имеются придаточные половые железы (передняя часть почки, становящаяся лейдиговым органом); выделения железы примешиваются к сперме.
· У некоторых рыб конец семяпровода расширен и образует семенной пузырек (не гомологичен органам того же названия у высших позвоночных).
· Известно о железистой функции семенного пузырька у некоторых представителей костистых рыб. От внутренних стенок семенника отходят внутрь семенные канальцы, сходящиеся к выводному протоку. По расположению канальцев семенники костистых рыб разделяют на две группы: циприноидные, или ацинозные, – у карповых, сельдевых, лососевых, сомовых, щуковых, осетровых, тресковых и др. ; перкоидные, или радиальные, – у окуневых, колюшковых и др. (рис. 24).
· В семенниках циприноидного типа семенные канальцы извиваются в различных плоскостях и без определённой системы. Вследствие этого на поперечных гистологических срезах видны их отдельные участки неправильной формы (так называемые ампулы). Выводной проток помещается в верхней части семенника. Края семенника округлы.
· В семенниках перкоидного типа семенные канальцы тянутся от стенок семенника радиально. Они прямые, выводной проток расположен в центре семенника. Семенник на поперечном срезе имеет треугольную форму.
· По стенкам канальцев (ампул) лежат крупные клетки – исходные семенные клетки, первичные сперматогонии, будущие сперматозоиды.
· Половые клетки появляются на ранних этапах развития у эмбрионов в генитальных складках, которые тянутся вдоль полости тела. У молоди лососей (горбуша, кета, нерка, сима, кижуч и атлантический лосось) первичные половые клетки обнаруживаются на стадии формирования первично-почечных протоков. У зародыша атлантического лосося первичные половые клетки были выявлены в возрасте 26 cyт. У мальков рыб уже можно найти половые железы в виде волосовидных тяжей.
· Овогонии – будущие икринки – образуются в результате деления зачатковых клеток зародышевого эпителия, это округлые, очень мелкие, не видимые простым глазом клетки. После овогониальных делений овогония превращается в овоцит. В дальнейшем в ходе овогенеза – развития яйцевых клеток – различают три периода: период синаптенного пути, период роста (малого – протоплазматического и большого – трофоплазматического) и период созревания.
· Каждый из этих периодов подразделяется на ряд фаз. Период синаптенного пути характеризуется главным образом преобразованием ядра клетки (овоцита). Затем наступает период малого –протоплазматического – роста, когда увеличение размеров овоцита происходит за счет накопления цитоплазмы. Здесь в развитии овоцитов выделяют ювенильную фазу и фазу однослойного фолликула.
· В ювенильной фазе овоциты ещё относительно небольшие, чаще всего округлой формы, с тонкой, бесструктурной, так называемой первичной (вырабатываемой самой яйцеклеткой) оболочкой, к которой прилегают отдельные фолликулярные клетки, а снаружи – клетки соединительной ткани. Ядро овоцита имеет хорошо заметную тонкую оболочку; округлое большое, оно почти всегда лежит в центре. По периферии ядра располагаются многочисленные ядрышки, в большинстве прилегающие к оболочке. В фазе однослойного фолликула собственная оболочка становится толще, над ней формируется фолликулярная оболочка с прилегающими отдельными соединительнотканными клетками.
· В этой же фазе в овоците часто можно обнаружить вителлогенную зону. Эта зона имеет ячеистую, как бы пенистую структуру и возникает в цитоплазме вокруг ядра, на некотором расстоянии от него (циркумнуклеарная зона). К концу фазы (и периода) овоциты увеличиваются настолько, что их можно различить с помощью лупы или даже невооруженным глазом.
· На протяжении формирования яйцевой клетки наряду с преобразованиями ядра в ней образуются и накапливаются питательные вещества, концентрирующиеся в желтке (белки и липиды) и чисто липидных включениях, которые затем, в период развития эмбриона, используются для его пластических и энергетических нужд. Этот процесс начинается в периоде большого роста овоцита, когда на периферии его появляются вакуоли, содержащие углеводы. Таким образом, период большого (трофоплазматического) роста овоцита характеризуется увеличением не только количества протоплазмы, но накоплением в нем питательных, трофических веществ – белка и жира.
· В период большого роста происходят вакуолизация цитоплазмы, появление желтка и наполнение им овоцита. Период большого роста также состоит из нескольких фаз. В фазе вакуолизации цитоплазмы овоциты, увеличенные по сравнению с предыдущей фазой, имеют несколько угловатую форму вследствие давления соседних клеток. Оболочки овоцита – собственная, фолликулярная, соединительнотканная – стали четче выраженными. На периферии овоцита образуются единичные мелкие вакуоли, которые, увеличиваясь в числе, создают более или менее густой слой. Это будущие кортикальные альвеолы, или гранулы. Содержимое вакуолей составляют углеводы (полисахариды), которые после оплодотворения яйца способствуют всасыванию под оболочку воды и образованию перивителлинового пространства. У некоторых видов (лосось, карп) раньше вакуолей в цитоплазме появляются жировые включения. В ядре ядрышки отходят от оболочки вглубь. В следующей фазе – первоначального накопления желтка – на периферии овоцита между вакуолями появляются отдельные мелкие шарики желтка, количество которых быстро растет, так что к концу фазы они занимают почти всю плазму овоцита.
· В собственной оболочке появляются тонкие канальцы, придающие ей радиальную исчерченность (Zona radiata); по ним в овоцит проникают питательные вещества. Над собственной оболочкой у некоторых рыб образуется ещё одна вторичная – оболочка (производная фолликулярных клеток, окружающих овоцит). Эта оболочка, разнообразная по структуре (студенистая, сотовая или ворсинчатая), после выхода овоцита из фолликула служит для прикрепления икринки к субстрату. Фолликулярная оболочка становится двуслойной. Границы ядра отчетливы, но стали извилистыми, “лапчатыми”.
· Следующая фаза – наполнение овоцита желтком – характеризуется очень сильным увеличением объёма желтка, частицы которого приобретают вместо шаровидной многогранную, глыбковидную форму. Вакуоли отжимаются к поверхности овоцита.
· Из-за преобладания в это время количественных изменений (без существенных морфологических сдвигов) некоторые исследователи считают нецелесообразным выделять эту фазу как самостоятельную. К концу фазы овоцит достигает дефинитивных размеров. Заметны изменения желтка и ядра: ядро начинает смещаться (к анимальному полюсу), его контуры становятся менее ясными; частицы желтка начинают сливаться. Заканчивается формирование вторичной оболочки.
· Последняя фаза развития – фаза зрелого овоцита. Частицы желтка у большинства рыб (за исключением вьюна, макропода, некоторых карповых) сливаются в гомогенную массу, овоцит становится прозрачным, цитоплазма сосредоточивается на периферии овоцита, ядро теряет контуры. Преобразования ядра вступают в завершающую стадию.
· Одно за другим следуют два деления созревания. В результате образуются ядро зрелого овоцита с гаплоидным числом хромосом и три редукционных тельца, которые не участвуют в дальнейшем развитии, отделяются от яйца и дегенерируют. После второго деления созревания митотическое развитие ядра доходит до метафазы и в этом состоянии остается до оплодотворения.
· Дальнейшее развитие (формирование женского пронуклеуса и отделение полярного тельца) происходит уже после оплодотворения.
· Сквозь собственную (Z. radiata) и студенистую оболочку проходит канал (микропиле), через который сперматозоид проникает в яйцеклетку при оплодотворении. У костистых рыб одно микропиле, у осетровых – несколько: у севрюги – до 13, белуги – до 33, черноморско-азовского осетра – до 52. Поэтому полиспермия возможна только у осетровых рыб, но не у костистых. При овуляции фолликулярная и соединительнотканная оболочки лопаются и остаются на яйценесущих пластинках, а освобожденный из них овоцит, окруженный собственной и студенистой оболочками, выпадает в полость яичника или полость тела. Здесь овулировавшие яйца находятся в полостной (овариальной) жидкости, сохраняя относительно длительное время способность к оплодотворению (табл. 3). В воде или вне полостной жидкости они эту способность быстро теряют.
· У акул и скатов, которым свойственно внутреннее оплодотворение, оплодотворенное яйцо, продвигаясь по половым путям, окружается ещё одной – третичной – оболочкой. Рогоподобное вещество этой оболочки образует твердую капсулу, надежно защищающую зародыш во внешней среде (см. рис. 34).
· В процессе развития овоцитов наряду с другими изменениями происходит колоссальное увеличение его размеров: так, по сравнению с овогониями, образованными при последнем овогониальном делении, объём зрелого овоцита возрастает у окуня в 1 049 440, у воблы – в 1 271 400 раз.
Таблица 3 Сохранение яйцами способности к оплодотворению (по Гинзбург, 1968, с сокращениями)
| Вид | t°C |
Время сохранения способности к оплодотворению
|
||
| Белуга Huso huso | 12-13,5 |
|
||
| Озерная форель Salmo truttamorpha lacustris | 1,7-8,9 10-12,5 0,4-1,0 |
|
||
| Щука Esox lucius | 3,5 10 24 |
|
||
| Золотая рыбка Carassius auratus | 4-10 |
|
||
| Судак Lucioperca lucioperca | 4-10 |
|
· У одной самки овоциты (а после овуляции – икринки) неодинаковы по величине: самые крупные могут превосходить самые мелкие в 1,5–2 раза. Это зависит от их расположения на яйценосной пластинке: овоциты, лежащие вблизи кровеносных сосудов, лучше снабжаются питательными веществами и достигают больших размеров.
· Отличительной особенностью процесса развития сперматозоидов – сперматогенеза – является многократное уменьшение клеток. Каждый исходный сперматогоний делится несколько раз, в результате чего возникает скопление сперматогониев под одной оболочкой, называемое цистой (стадия размножения). Образовавшийся при последнем делении сперматогоний несколько увеличивается, в его ядре происходят мейотические преобразования и сперматогоний превращается в сперматоцит I порядка (стадия роста). Затем наступают два последовательных деления (стадия созревания): сперматоцит I порядка делится на два сперматоцита II порядка, вследствие деления которых образуются две сперматиды. В следующей – завершающей – стадии формирования сперматиды превращаются в сперматозоиды. Таким образом, из каждого сперматоцита образуются четыре сперматиды с половинным (гаплоидным)набором хромосом. Оболочка цисты лопается, и сперматозоиды наполняют семенной каналец. Через семяпроводы созревшие сперматозоиды выходят из семенника, а затем по протоку – наружу.
· Характерными особенностями развития семенников является сильная неравномерность (асинхронность) развития органа в целом. Особенно сильно эта неравномерность проявляется у впервые созревающих рыб, но вполне отчетливо выражена и у нерестующих повторно половозрелых особей. В результате практически все самцы нерестуют порционно и в течение длительного периода от них можно получать сперму.
· Процесс созревания половых клеток у разных рыб проходит, в общем, по одной схеме. По мере развития половых клеток внутри яичников и семенников меняются и внешний вид, и размеры гонад. Это побудило составить так называемую шкалу зрелости гонад, пользуясь которой можно было бы по внешним признакам гонад определить степень зрелости половых продуктов, что очень важно в научных и промысловых исследованиях. Чаще других используют универсальную 6-балльную шкалу, в основе которой лежат общие признаки для разных видов рыб (табл. 4, 5; рис. 25, 26). Предложены также и другие шкалы, в которых учитывают особенности созревания определённых групп рыб. Так, для яичников карповых и окуневых В. М. Мейеном предложена 6-балльная шкала, а для семенников С. И. Кулаевым – 8-балльная.
· У подавляющего большинства рыб осеменение наружное. У хрящевых рыб, которым свойственны внутреннее осеменение и живорождение, имеются соответствующие изменения в строении полового аппарата. Развитие зародышей у них происходит в заднем отделе яйцеводов, получивших название матки. Из костистых рыб живорождение свойственно гамбузии, морскому окуню, многим аквариумным рыбкам. У них молодь развивается в яичнике.
Таблица 4 Шкала зрелости гонад. Самки ..
| Стадия | Внешний вид гонад | Микроскопическое строение |
| I ювенильная неполовозрелая (juvenis) | Половые железы в виде прозрачных тонких нитей. Невооруженным глазом пол различить нельзя | Половые клетки – овогонии – обнаруживаются среди клеток герминативного эпителия (овогониальный период) |
Стадия не повторяется (бывает один раз в жизни)
| II | Яичники представлены стекловидными тяжами; мягкие, розовато-желтоватого оттенка. Сквозь оболочку яичника видны невооруженным глазом или под лупой очень мелкие прозрачные овоциты. Яичник кажется зернистым. По стенкам тянутся крупные кровеносные сосуды. Яйценесущие пластинки при разрезе стенок яичника отделимы друг от друга, видно их расположение | Многочисленны овоциты периода малого (протоплазматического) роста, старшая генерация которых находится в фазе однослойного фолликула. Они округлой или многоугольной формы, плотно прилегают друг к другу. Имеются половые клетки предыдущих фаз развития |
У неполовозрелых рыб эта стадия следует за I; в яичниках половозрелых самок II стадия наступает после того, как исчезают признаки прошедшего нереста, т. е. после VI стадии
| III | Яичники округлой формы, жёлтовато-оранжевого цвета, занимают около 1/3– 1/2 длины полости тела. Они наполнены мелкими непрозрачными желтоватыми или беловатыми икринками, хорошо видными невооруженным глазом. При разрезе яичника икринки держатся комками; яйценесущие пластинки ещё видны. По стенкам яичника проходят крупные ветвящиеся кровеносные сосуды | Овоциты лежат более густо вследствие увеличения их размеров. Они находятся в начале периода большого (трофоплазматического) роста: основная масса овоцитов проходит фазы ва куолизации цитоплазмы и начала желткообразования. Имеются младшие генерации. У уже нерестовавших самок могут встретиться резорбирующиеся невыметанные икринки |
| IV | Яичники сильно увеличены в объёме и занимают больше половины – иногда до 2/3 полости тела. Они светло-оранжевого цвета, туго набиты непрозрачными икринками. Стенки яичника прозрачны. При разрезе их выпадают отдельные икринки. Яйценесущие пластинки неразличимы. Макроскопически легко заметить переход овоцитов старшей генерации в следующую фазу: в яичнике, близком к зрелости, среди желтых мутных овоцитов появляются одиночные более крупные и прозрачные икринки. Количество таких икринок увеличивается. | Овоциты старшей генерации находятся в конце периода трофоплазматического роста, т. е. в фазе наполнения желтком. Имеются овоциты младших генераций. Иногда встречаются остатки дегенерирующих зрелых икринок (у половозрелых рыб) |
| V | Яичники достигают максималь ных размеров, они наполнены икринками, вытекающими при слабом поглаживании брюшка (а после гипофизарных инъекций – и без какого-либо надавливания). Овулировавшие икринки прозрачны, шаровидны | Овоциты старшей генерации достигли дефинитивных размеров. Глыбки желтка сливаются (у большинства видов). Ядро неразличимо. Овоциты выходят из фолликулов. Присутствуют овоциты младших генераций |
| VI | Выбой, яичник после нереста. Стенки яичника спадаются, становятся дряблыми, непрозрачными, складчатыми, красновато-синеватого цвета. Опустошенный яичник сильно уменьшается в объёме | Опустевшие фолликулы, дегенерирующие оставшиеся невыметанными зрелые икринки, овоциты молодых генерации |
Через некоторое время воспаление проходит, яичник постепенно светлеет, становится светло-розовым и переходит в стадию II.
Таблица 5 Шкала зрелости гонад. Самцы .
Стадия не повторяется
У многократно нерестующих рыб железа переходит затем во II стадию |
Нервная система и органы чувств
· Нервная система рыб представлена центральной нервной системой и связанной с ней периферической и вегетативной (симпатической) нервной системой. Центральная нервная система состоит из головного и спинного мозга. К периферической нервной системе относятся нервы, отходящие от головного и спинного мозга к органам. Вегетативная нервная система в основе имеет многочисленные ганглии и нервы, иннервирующие, мышцы внутренних органов и кровеносных сосудов сердца. Нервная система рыб по сравнению с нервной системой высших позвоночных характеризуется рядом примитивных черт.
· Центральная нервная система имеет вид нервной трубки, тянущейся вдоль туловища; часть ее, лежащая над позвоночником и защищенная верхними дугами позвонков, образует спинной мозг, а расширенная передняя часть, окруженная хрящевым или костным черепом, составляет головной мозг. Трубка имеет внутри полость (невроцель), представленную в головном мозгу желудочками мозга. В толще мозга различают серое вещество, слагающееся из тел нервных клеток и коротких отростков (дендритов), и белое вещество, образованное длинными отростками нервных клеток – нейритами или аксонами.
· Общая масса мозга у рыб мала: она составляет в среднем у современных хрящевых рыб 0,06 – 0,44%, у костных – 0,02 – 0,94% от массы тела, в том числе у налима 1/700 массы тела, щуки 1/3000, акулы – 1/37000 , в то время как у летающих птиц и млекопитающих 0,2 – 8,0 и 6,3 – 3,0%.
· В строении головного мозга сохраняются примитивные черты: отделы мозга располагаются линейно. В нем выделяют передний мозг, промежуточный, средний, мозжечок и продолговатый, переходящий в спинной мозг (рис. 27). Полости переднего, промежуточного и продолговатого мозга называются желудочками: полость среднего мозга–сильвиевым водопроводом (она соединяет полости промежуточного и продолговатого мозга, т. е. третий и четвертый желудочки).
· Передний мозг благодаря продольной борозде имеет вид двух полушарий. К ним прилегают обонятельные луковицы (первичный обонятельный центр) или непосредственно (у большинства видов), или через обонятельный тракт (карповые, сомовые, тресковые).
· В крыше переднего мозга нет нервных клеток. Серое вещество в виде полосатых тел сосредоточено главным образом в основании и обонятельных долях, выстилает полость желудочков и составляет главную массу переднего мозга. Волокна обонятельного нерва связывают луковицу с клетками обонятельной капсулы.
· Передний мозг является центром обработки информации, поступающей от органов обоняния. Благодаря своей связи с промежуточным и средним мозгом он участвует в регуляции движения и поведения. В частности, передний мозг принимает участие в формировании способности к таким актам, как икрометание, охрана икры, образование стаи и т. д.
· В промежуточном мозге развиты зрительные бугры. От них отходят зрительные нервы, образующие хиазму (перекрест, т. е. часть волокон правого нерва переходит в левый нерв и наоборот). На нижней стороне промежуточного мозга (гипоталамус) имеется воронка, к которой прилегает гипофиз, или питуитарная железа; в верхней части промежуточного мозга развивается эпифиз, или пинеальная железа. Гипофиз и эпифиз являются железами внутренней секреции.
· Промежуточный мозг выполняет многочисленные функции. Он воспринимает раздражения от сетчатки глаза, участвует в координации движений, в переработке информации от других органов чувств. Гипофиз и эпифиз осуществляют гормональную регуляцию обменных процессов. Средний мозг наибольший по объёму. Он имеет вид двух полушарий (зрительные доли). Зрительные доли являются первичными зрительными центрами, воспринимающими возбуждение. Из этих долей берут начало волокна зрительного нерва. В среднем мозгу обрабатываются сигналы, идущие от органов зрения и равновесия; здесь помещаются центры связи с мозжечком, продолговатым и спинным мозгом.
· Мозжечок расположен в задней части мозга и может иметь форму или маленького бугорка, прилегающего сзади к среднему мозгу, или большого мешковидно-вытянутого образования, примыкающего сверху к продолговатому мозгу. Особенно большого развития достигает мозжечок у сомов, а у мормируса относительная его величиная вляется наибольшей среди остальных позвоночных. В мозжечке рыб, как и высших позвоночных, имеются клетки Пуркинье. Мозжечок является центром всех моторных иннервации при плаваниии, схватывании пищи. Он обеспечивает координацию движений, поддержание равновесия, мышечную деятельность, связан с рецепторами органов боковой линии.
· Пятый отдел головного мозга, продолговатый мозг, без резкой границы переходит в спинной мозг. Полость продолговатого мозга –четвертый желудочек – продолжается в полость спинного мозга – невроцель. Значительная масса продолговатого мозга состоит из белого вещества.
· От продолговатого мозга отходит большая часть (шесть из десяти) черепно-мозговых нервов. Он является центром регуляции деятельности спинного мозга и вегетативной нервной системы. В нем располагаются наиболее важные жизненные центры, регулирующие деятельность дыхательной, скелетно-мышечной, кровеносной, пищеварительной, выделительной систем, органов слуха и равновесия, вкуса, боковой линии, электрических органов у имеющих их рыб и т. д. Поэтому при разрушении продолговатого мозга, например при перерезке туловища позади головы, наступает быстрая смерть рыбы. Через приходящие в продолговатый мозг спинномозговые волокна осуществляется связь продолговатого и спинного мозга.
· От головного мозга отходит 10 пар черепно-мозговых нервов:
· I – обонятельный нерв (nervus olfactorius) – от чувствующего эпителия обонятельной капсулы доводит раздражения до обонятельных луковиц переднего мозга;
· II – зрительный нерв (n. opticus) – тянется до сетчатки глаза от зрительных бугров промежуточного мозга;
· III – глазодвигательный нерв (n. oculomotorius) – иннервирует мышцы глаза, отходя от среднего мозга;
· IV – блоковый нерв (n. trochlearis), глазодвигательный, тянущийся от среднего мозга кодной из мышц глаза;
· V – тройничный нерв (n. trigeminus), отходящийот боковой поверхности продолговатого мозга и дающий три основные ветви: глазничную, верхнечелюстную и нижнечелюстную;
· VI –отводящий нерв (n. abducens) – тянется от дна мозга к прямой мышце глаза;
· VII – лицевой нерв (n. facialis) – отходит от продолговатого мозга и дает многочисленные разветвления к мускулатуре подъязычной дуги, слизистой ротовой полости, коже головы (в том числе боковой линии головы);
· VIII – слуховой нерв (n. acusticus) –связывает продолговатый мозг и слуховой аппарат;
· IX – языкоглоточный нерв (n. glossopharingeus) – идет от продолговатого мозга к глотке, иннервирует слизистую глотки и мускулатуру первой жаберной дуги;
· X – блуждающий нерв (n. vagus) – наиболее длинный. Связывает продолговатый мозг с жаберным аппаратом, кишечным трактом, сердцем, плавательным пузырем, боковой линией.
· Степень развития разных отделов головного мозга различна у разных групп рыб и связана с образом жизни.
· Передний мозг (и обонятельные доли) относительно сильнее развит у хрящевых рыб (акулы и скаты) и слабее – у костистых. У малоподвижных, например донных, рыб мозжечок мал, но сильнее развиты передний и продолговатый отделы мозга в соответствии с большой ролью обоняния и осязания в их жизни (камбалы). У хорошо плавающих рыб (пелагических, питающихся планктоном, или хищничающих), наоборот, гораздо большее развитие получают средний мозг (зрительные доли) и мозжечок (в связи с необходимостью быстрой координации движения). Рыбы, обитающие в мутной воде, имеют маленькие зрительные доли, небольшой мозжечок. Слабо развиты зрительные доли у глубоководных и слепых рыб.
· Спинной мозг является продолжением продолговатого мозга. Он имеет форму округлого тяжа и лежит в канале, образованном верхними дугами позвонков. В спинном мозге серое вещество расположено внутри, а белое–снаружи. От спинного мозга метамерно, соответственно каждому позвонку, отходят спинномозговые нервы, иннервирующие поверхность тела, туловищные мышцы, а благодаря соединению спинномозговых нервов с ганглиями симпатической нервной системы – и внутренние органы.
· Вегетативная нервная система у хрящевых рыб представлена разобщенными ганглиями, лежащими вдоль позвоночника. Клетки ганглиев своими отростками контактируют со спинномозговыми нервами и внутренними органами.
· У костистых рыб ганглии вегетативной нервной системы соединяются двумя продольными нервными стволами. Соединительные ветви ганглиев связывают вегетативную нервную систему с центральной. Взаимосвязи центральной и вегетативной нервной систем создают возможность некоторой взаимозаменяемости нервных центров.
· Вегетативная нервная система действует в определённой степени автономно, независимо от центральной нервной системы и определяет непроизвольную, автоматическую деятельность внутренних органов даже в том случае, если ее связь с центральной нервной системой нарушена.
· Реакцию организма рыбы на внешние и внутренние раздражения определяет рефлекс. У рыб можно выработать условный рефлекс на свет, форму, запах, вкус, звук. По сравнению с высшими позвоночными у рыб условные рефлексы образуются медленнее, а гаснут быстрее. Тем не менее и аквариумные, и прудовые рыбы вскоре после начала регулярного кормления скапливаются в определённое время у кормушек. Привыкают они и к звукам во время кормления (постукивание по стенкам аквариума, звон колокольчика, свист, удары) и какое-то время подплывают на эти раздражители и при отсутствии пищи.
· Органы восприятия окружающей среды (органы чувств) рыб обладают рядом особенностей, отражающих их приспособленность к условиям жизни.
· Способность рыб воспринимать информацию из окружающей среды многообразна. Их рецепторы могут улавливать различные раздражения как физической, так и химической природы: давление, звук, цвет, температуру, электрические и магнитные поля, запах, вкус.
· Одни раздражения воспринимаются в результате непосредственного прикосновения (осязание, вкус), другие–на расстоянии, дистанционно.
· Органы, воспринимающие химические, тактильные (прикосновение) , электромагнитные, температурные и другие раздражения, имеют простое строение. Раздражения улавливаются свободными нервными окончаниями чувствующих нервов на поверхности кожи. У некоторых групп рыб они представлены специальными органами или входят в состав боковой линии.
· В связи с особенностями жизненной среды у рыб большое значение имеют системы химического чувства. Химические раздражения воспринимаются при помощи обоняния (ощущения запаха) или при помощи органов необонятельной рецепции, обеспечивающих восприятие вкуса, изменения активности среды и т. д. Химическое чувство называется хеморецепцией, а чувствующие органы – хеморецепторами.
· Органы обоняния. У рыб, как и у других позвоночных, они находится в передней части головы и представлены парными обонятельными (носовыми) мешками (капсулами), открывающимися наружу отверстиями-ноздрями. Дно носовой капсулы выстлано складками эпителия, состоящего из опорных и чувствующих клеток (рецепторов) . Наружная поверхность чувствующей клетки снабжена ресничками, а основание связано с окончаниями обонятельного нерва. В обонятельном эпителии многочисленны клетки, секретирующие слизь.
· Ноздри расположены у хрящевых рыб на нижней стороне рыла впереди рта, у костистых – на дорсальной стороне между ртом и глазами. Круглоротые имеют по одной ноздре, настоящие рыбы –по две. Каждая ноздря разделяется кожистой перегородкой на два отверстия. Вода проникает в переднее из них, омывает полость и выходит через заднее отверстие, омывая и раздражая при этом волоски рецепторов. Под влиянием пахучих веществ в обонятельном эпителии происходят сложные процессы: перемещения липидов, белково-мукополисахаридных комплексов и кислой фосфатазы.
· Величина ноздрей связана с образом жизни рыб: у подвижных рыб они небольшие, так как при быстром плавании вода в обонятельной полости обновляется быстро; у рыб малоподвижных, наоборот, ноздри большие, они пропускают через носовую полость больший объём воды, что особенно важно для плохих пловцов, в частности обитающих у дна.
· Рыбы обладают тонким обонянием, т. е. пороги обонятельной чувствительности у них очень низки. Это особенно относится к ночными сумеречным рыбам, а также к живущим в мутных водах, которым зрение мало помогает в отыскании пищи и общении с сородичами. Наиболее удивительна чувствительность обоняния у проходных рыб. Дальневосточные лососи совершенно точно находят путь от мест нагула в море к нерестилищам в верховьях рек, где они вывелись несколько лет назад. При этом они преодолевают огромные расстояния и препятствия – течения, пороги, перекаты. Однако рыбы верно проходят путь лишь в том случае, если у них открыты ноздри; если же обоняние выключено (ноздри заполнены ватой или вазелином), то рыбы идут беспорядочно. Предполагают, что лососи в начале миграции ориентируются по солнцу и примерно за 800 км от родной реки безошибочно определяют путь благодаря хеморецепции.
· В опытах при омывании носовой полости этих рыб водой с родного нерестилища в обонятельной луковице мозга возникала сильная электрическая реакция. На воду из нижерасположенных притоков реакция была слабой, а на воду с чужих нерестилищ рецепторы вообще не реагировали.
· Молодь нерки Oncorhynchus nerka может различать при помощи клеток обонятельной луковицы воду разных озер, растворы различных аминокислот в разведении 10-4, а также концентрацию кальция в воде. Не менее поразительна аналогичная способность европейского угря, мигрирующего из Европы к нерестилищам, расположенным в Саргассовом море. Подсчитано, что угорь в состоянии распознавать концентрацию, создаваемую разведением 1 г фенилэтилового спирта в соотношении 1: 3 • 10-18. Высокая избирательная чувствительность к гистамину обнаружена у карпа.
· Обонятельный рецептор рыб кроме химических способен воспринимать механические воздействия (струи потока) и изменения температуры.
· Органы вкуса. Они представлены вкусовыми почками, образованными скоплениями чувствующих (и опорных) клеток. Основания чувствующих клеток оплетены концевыми разветвлениями лицевого, блуждающего и языкоглоточного нервов.
· Восприятие химических раздражителей осуществляется также свободными нервными окончаниями тройничного, блуждающего и спинномозговых нервов. Восприятие вкуса рыбами не обязательно связано с ротовой полостью, так как вкусовые почки расположены как в слизистой ротовой полости и на губах, так и в глотке, на усиках, жаберных лепестках, плавниковых лучах и по всей поверхности тела, в том числе на хвосте.
· Сом воспринимает вкус главным образом при помощи усов: именно в их эпидермисе сосредоточены скопления вкусовых почек. У одной и той же особи количество вкусовых почек увеличивается по мере увеличения размеров тела. Рыбы различают вкусовые особенности пищи: горькое, соленое, кислое, сладкое. В частности, восприятие солености связано с ямковидным органом, помещающимся в ротовой полости.
· Чувствительность органов вкуса у некоторых рыб очень высока: например, пещерные рыбы Anoptichthys, будучи слепыми, ощущают раствор глюкозы в концентрации 0,005%.
· Органы чувств боковой линии. Специфическим органом, свойственным только рыбам и живущим в воде амфибиям, является орган бокового чувства, или боковой линии. Это сейсмосенсорные специализированные кожные органы. Наиболее просто органы боковой линии устроены у круглоротых и личинок карповых. Чувствующие клетки (механорецепторы) лежат среди скоплений эктодермальных клеток на поверхности кожи или в мелких ямках. У основания они оплетены конечными разветвлениями блуждающего нерва, а на участке, возвышающемся над поверхностью, имеют реснички, воспринимающие колебания воды. У большинства взрослых костистых эти органы представляют собой погруженные в кожу каналы, тянущиеся по бокам тела вдоль средней линии. Канал открывается наружу через отверстия (поры) в чешуйках, расположенных над ним (рис. 28).
· Разветвления боковой линии имеются и на голове. На дне канала (группами лежат чувствующие клетки с ресничками. Каждая такая группа рецепторных клеток вместе с контактирующими с ними нервными волокнами образует собственно орган – невромаст. Вода свободно протекает через канал, и реснички ощущают её давление. При этом возникают нервные импульсы разной частоты. Органы боковой линии связаны с центральной нервной системой блуждающим нервом.
· Боковая линия может быть полной, т. е. тянуться по всей длине тела, или неполной и даже отсутствовать, но в последнем случае сильно развиваются головные каналы (у сельдей). Боковая линия дает возможность рыбе ощущать изменение давления текущей воды, вибрации (колебания) низкой частоты, инфразвуковые колебания, а многим рыбам – и электромагнитные поля. Боковая линия улавливает давление струящегося, движущегося потока, изменения давления с погружением на глубину она не воспринимает. Улавливая колебания водной толщи, органы боковой линии дают возможность рыбе обнаруживать поверхностные волны, течения, подводные неподвижные предметы (скалы, рифы) и движущиеся предметы (враги, добыча), плавать днем и ночью, в мутной воде и даже будучи ослепленной. Это весьма чувствительный орган: проходные рыбы ощущают им в море даже очень слабые токи пресной речной воды.
· Способность улавливать отраженные от живых и неживых объектов волны очень важна для глубоководных рыб, так как в темноте больших глубин невозможно обычное зрительное восприятие окружающих предметов, общение между особями.
· Предполагают, что волны, создающиеся во время брачных игр многих рыб, воспринимаемые боковой линией самки или самца, служат для них сигналом. Функцию кожного чувства выполняют итак называемые кожные почки – клетки, имеющиеся в покровах головы и усиков, к которым подходят нервные окончания, однако они имеют гораздо меньшее значение.
· Органы осязания. Органами осязания служат скопления чувствующих клеток (осязательные тельца), разбросанные по поверхности тела. Они воспринимают прикосновение твердых предметов (тактильные ощущения), давление воды, а также изменение температуры (тепло–холод) и боль.
· Особенно много чувствующих кожных почек находится во рту и на губах. У некоторых рыб функцию органов осязания выполняют удлиненные лучи плавников: у гурами это первый луч брюшного плавника, у триглы (морской петух) осязание связано с лучами грудных плавников, ощупывающими дно, и т. д. У обитателей мутных вод или донных рыб, наиболее активных ночью, наибольшее количество чувствующих почек сосредоточено на усиках и плавниках. Однако у сомов усы служат рецепторами вкуса, а не осязания.
· Механические травмы и боль рыбы, по-видимому, ощущают слабее, чем другие позвоночные: акулы, набросившиеся на добычу, не реагируют на удары острым предметом в голову; при операциях рыбы бывают часто относительно спокойны и т. д.
· Терморецепторы. Ими являются находящиеся в поверхностных слоях кожи свободные окончания чувствующих нервов, при помощи которых рыбы воспринимают температуру воды. Различают рецепторы, воспринимающие тепло (тепловые) и холод (холодовые). Точки восприятия тепла найдены, например, у щуки на голове, восприятия холода – на поверхности тела. Костистые рыбы улавливают перепады температуры в 0,1–0,4°С.
· Органы электрического чувства. Органы восприятия электрического и магнитного полей располагаются в коже на всей поверхности тела рыб, но главным образом в разных участках головы и вокруг нее. Они сходны с органами боковой линии – это ямки, заполненные слизистой массой, хорошо проводящей ток; на дне ямок помещаются чувствующие клетки (электрорецепторы), передающие нервные импульсы в мозг. Иногда они входят в состав системы боковой линии. Электрическими рецепторами у хрящевых рыб служат и ампулы Лоренцини. Анализ информации, получаемой электрорецепторами, осуществляет анализатор боковой линии (в про долговатом мозгу и мозжечке). Чувствительность рыб к току велика – до 1 мкВ/см2. Предполагают, что восприятие изменения электромагнитного поля Земли позволяет рыбам обнаруживать приближение землетрясения за 6–8 и даже за 22–24 ч до начала, в радиусе до 2 тыс. км.
· Органы зрения. Органы зрения рыб устроены в основном так же, как у других позвоночных. Сходен с остальными позвоночными у них и механизм восприятия зрительных ощущений: свет проходит в глаз через прозрачную роговицу, далее зрачок – отверстие в радужной оболочке – пропускает его на хрусталик, а хрусталик передает фокусирует свет на внутреннюю стенку глаза сетчатку, где и происходит его непосредственное восприятие (рис. 29). Сетчатка состоит из светочувствительных (фоторецепторные), нервных, а также опорных клеток.
· Светочувствительные клетки располагаются со стороны пигментной оболочки. В их отростках, имеющих форму палочек и колбочек, имеется светочувствительный пигмент. Количество этих фоторецепторных клеток очень велико –на 1 мм2 сетчатки у карпа их насчитывается 50 тыс. (у кальмара –162 тыс. , паука–16 тыс. , человека – 400 тыс. , совы – 680 тыс. ). Посредством сложной системы контактов конечных разветвлений чувствующих клеток и дендритов нервных клеток световые раздражения поступают в зрительный нерв.
· Колбочки при ярком свете воспринимают детали предметов и цвет. Палочки воспринимают слабый свет, но детального изображения создать не могут.
· Положение и взаимодействие клеток пигментной оболочки, палочек и колбочек меняются в зависимости от освещенности. На свету пигментные клетки расширяются и прикрывают находящиеся около них палочки; колбочки подтягиваются к ядрам клеток и таким образом передвигаются к свету. В темноте к ядрам подтягиваются палочки (и оказываются ближе к поверхности); колбочки приближаются к пигментному слою, а сократившиеся в темноте пигментные клетки прикрывают их (рис. 30).
· Количество рецепторов разного рода зависит от образа жизни рыб. У дневных рыб в сетчатке превалируют колбочки, у сумеречных и ночных – палочки: у налима палочек в 14 раз больше, чем у щуки. У глубоководных рыб, живущих в темноте глубин, колбочек нет, а палочки становятся больше и количество их резко увеличивается – до 25 млн/мм2 сетчатки; вероятность улавливания даже слабого света возрастает. Большая часть рыб различает цвета, что подтверждается возможностью выработки у них условных рефлексов на определённый цвет – синий, зеленый, красный, жёлтый, голубой.
· Некоторые отступления от общей схемы строения глаза рыбы связаны с особенностями жизни в воде. Глаз рыбы эллипсовидный. В числе других он имеет серебристую оболочку (между сосудистой и белковой), богатую кристалликами гуанина, которая придает глазу зеленовато-золотистый блеск.
· Роговица почти плоская (а не выпуклая), хрусталик шаровидный (а не двояковыпуклый) – это расширяет поле зрения. Отверстие в радужной оболочке – зрачок – может изменять диаметр только в небольших пределах. Век у рыб, как правило, нет. Лишь акулы имеют мигательную перепонку, закрывающую глаз как занавеска, и некоторые сельди и кефали – жировое веко – прозрачную пленку, закрывающую часть глаза.
· Расположение глаз по бокам головы (у большинства видов) является причиной того, что рыбы обладают в основном монокулярным зрением, а способность к бинокулярному зрению весьма ограничена. Шаровидность хрусталика и перемещение его вперед к роговице обеспечивает широту поля зрения: свет в глаз попадает со всех сторон. Угол зрения по вертикали составляет 150°, по горизонтали– 168–170°. Но вместе с тем шаровидность хрусталика обусловливает близорукость рыб. Дальность их зрения ограничена и колеблется в связи с мутностью воды от нескольких сантиметров до нескольких десятков метров.
· Видение на дальние расстояния становится возможным благодаря тому, что хрусталик может быть оттянут специальной мышцей–серповидным отростком, идущим от сосудистой оболочки дна глазного бокала.
· При помощи зрения рыбы ориентируются и относительно предметов, находящихся на земле. Улучшение зрения в темноте достигается наличием отражательного слоя (тапетум) – кристалликов гуанина, подстилаемых пигментом. Этот слой не пропускает свет к лежащим позади сетчатки тканям, а отражает его и возвращает вторично на сетчатку. Так увеличивается возможность рецепторов использовать свет, попавший в глаз.
· В связи с условиями обитания глаза рыб могут сильно видоизменяться. У пещерных или абиссальных (глубоководных) форм глаза могут редуцироваться и даже исчезать. Некоторые же глубоководные рыбы, наоборот, имеют огромные глаза, позволяющие улавливать совсем слабые следы света, или телескопические глаза, собирающие линзы которых рыба может поставить параллельно и обрести бинокулярное зрение. Глаза некоторых угрей и личинок ряда тропических рыб вынесены вперед на длинных выростах (стебельчатые глаза).
· Необычна модификация глаз у четырехглазки из Центральной и Южной Америки. Ее глаза помещаются на верху головы, каждый из них разделен перегородкой на две самостоятельные части: верхней рыба видит в воздухе, нижней– в воде. В воздушной среде могут функционировать глаза рыб, выползающих на берег или деревья.
· Роль зрения как источника информации из внешнего мира для большинства рыб очень велика: при ориентации во время движения, при отыскивании и захвате пищи, при сохранении стаи, в нерестовый период (восприятие оборонительных и агрессивных поз и движений самцами-соперниками, а между особями разных полов – брачного наряда и нерестового “церемониала”), в отношениях жертва –хищник и т. д.
· Способность рыб воспринимать свет издавна использовалась в рыболовстве (лов рыбы на свет факела, костра и т. д. ).
· Известно, что рыбы разных видов неодинаково реагируют на свет разной интенсивности и разной длины волны, т. е. разного цвета. Так, яркий искусственный свет привлекает одних рыб (каспийская килька, сайра, ставрида, скумбрия и др. ) и отпугивает других (кефаль, минога, угорь и т. д. ). Так же избирательно относятся разные виды к разным цветам и разным источникам света – надводным и подводным. Все это положено в основу организации промышленного лова рыбы на электросвет (так ловят кильку, сайру и других рыб).
· Орган слуха и равновесия рыб. Он расположен в задней части черепной коробки и представлен лабиринтом; ушных отверстий, ушной раковины и улитки нет, т. е. орган слуха представлен внутренним ухом. Наибольшей сложности достигает он у настоящих рыб: большой перепончатый лабиринт помещается в хрящевой или костной камере под прикрытием ушных костей. В нем различают верхнюю часть – овальный мешочек (ушко, utriculus) и нижнюю – круглый мешочек (sacculus). От верхней части во взаимно перпендикулярных направлениях отходят три полукружных канала, каждый из которых на одном конце расширен в ампулу (рис. 31). Овальный мешочек с полукружными каналами составляет орган равновесия (вестибулярный аппарат). Боковое расширение нижней части круглого мешочка (lagena), являющееся зачатком улитки, не получает у рыб дальнейшего развития. От круглого мешочка отходит внутренний лимфатический (эндолимфатический) канал, который у акул и скатов через специальное отверстие в черепе выходит наружу, а у остальных рыб слепо заканчивается у кожи головы.
· Эпителий, выстилающий отделы лабиринта, имеет чувствующие клетки с волосками, отходящими во внутреннюю полость. Основания их оплетены разветвлениями слухового нерва. Полость лабиринта заполнена эндолимфой, в ней находятся “слуховые” камешки, состоящие из углекислой извести (отолиты), по три с каждой стороны головы: в овальном и круглом мешочке и лагене. На отолитах, как и на чешуе, образуются концентрические слои, поэтому отолиты, и особенно наибольший, часто используют для определения возраста рыб, а иногда и для систематических определений, так как их размеры и контуры неодинаковы у различных видов.
· У большинства рыб наибольший отолит располагается в круглом мешочке, но у карповых и некоторых других – в лагене,
· С лабиринтом связано чувство равновесия: при передвижении рыбы давление эндолимфы в полукружных каналах, а также со стороны отолита изменяется и возникшее раздражение улавливается нервными окончаниями. При экспериментальном разрушении верхней части лабиринта с полукружными каналами рыба теряет способность удерживать равновесие и лежит на боку, спине или брюхе. Разрушение нижней части лабиринта не ведет к утрате равновесия.
· С нижней частью лабиринта связано восприятие звуков: при удалении нижней части лабиринта с круглым мешочком и лагеной рыбы не в состоянии различать звуковые тона (при попытках выработать условный рефлекс). В то же время рыбы без овального мешочка и полукружных каналов, т. е. без верхней части лабиринта, дрессировке поддаются. Таким образом, было показано, что рецепторами звука являются именно круглый мешочек и лагена.
· Рыбы воспринимают как механические, так и звуковые колебания: частотой от 5 до 25 Гц – органами боковой линии, от 16 до 13 000 Гц – лабиринтом. Некоторые виды рыб улавливают колебания, находящиеся на границе инфразвуковых волн и боковой линией, и лабиринтом.
· Острота слуха у рыб ниже, чем у высших позвоночных, и у разных видов неодинакова: язь воспринимает колебания, длина волны которых составляет 25–5524 Гц, серебряный карась – 25–3840, угорь – 36–650 Гц, причем низкие звуки улавливаются ими лучше.
· Рыбы улавливают и те звуки, источник которых находится не в воде, а в атмосфере, несмотря на то что такой звук на 99,9% отражается поверхностью воды и, следовательно, в воду проникает только 0,1 % образующихся звуковых волн. В восприятии звука у карповых, сомовых рыб большую роль играет плавательный пузырь, соединенный с лабиринтом и служащий резонатором.
· Рыбы могут и сами издавать звуки. Звукоиздающие органы у рыб различны: плавательный пузырь (горбыли, губаны и др. ), лучи грудных плавников в комбинации с костями плечевого пояса (сомы), челюстные и глоточные зубы (окуневые и карповые) и др. В связи с этим неодинаков и характер звуков: они могут напоминать удары, цоканье, свист, ворчанье, хрюканье, писк, кваканье, рычанье, треск, рокот, звон, хрип, гудок, крики птиц и стрекотанье насекомых. Сила и частота звуков, издаваемых рыбами одного вида, зависит от пола, возраста, пищевой активности, здоровья, причиняемой боли и т. д.
· Звучание и восприятие звуков имеет большое значение в жизнедеятельности рыб: оно помогает особям разного пола найти друг друга, сохранить стаю, сообщить сородичам о присутствии пищи, охранять территорию, гнездо и потомство от врагов, является стимулятором созревания во время брачных игр, т. е. служит важным средством общения. Предполагают, что у глубоководных рыб, рассредоточенных в темноте на океанических глубинах, именно слух в сочетании с органами боковой линии и обонянием обеспечивает общение, тем более что звукопроводимость, более высокая в воде, чем в воздухе, на глубине возрастает. Особенно важен слух для ночных рыб и обитателей мутных вод.
· Реакция разных рыб на посторонние звуки различна: при шуме одни уходят в сторону, другие толстолобик, семга, кефаль–выпрыгивают из воды. Это используют при организации лова рыбы (лов кефали рогожами, колокол, отпугивающий ее от ворот кошелькового невода, и т. д.). В период нереста карпа в рыбоводных хозяйствах запрещают проезд около нерестовых прудов, а в старину во время нереста леща запрещали колокольный звон.
Железы внутренней секреции
· Железами внутренней секреции являются гипофиз, эпифиз, надпочечники, поджелудочная, щитовидная и ультимобранхиальная (подпищеводная) железы. Они выделяют гормоны в кровь. Их деятельность регулируется центральной нервной системой.
· Наиболее изученными у рыб являются гипофиз и щитовидная железа. Гипофиз – непарное, неправильной овальной формы образование, отходящее от нижней стороны промежуточного мозга (гипоталамуса); очертания, размеры и положение его чрезвычайно разнообразны у рыб разных групп. У сазана, карпа, большинства других рыб гипофиз сердцевидной формы, лежит почти перпендикулярно мозгу; у серебряного карася он вытянут, немного сплющен с боков, лежит параллельно мозгу.
· В гипофизе различают два основных отдела различного происхождения: мозговой (нейрогипофиз), составляющий внутреннюю часть железы, который развивается из нижней стенки промежуточного мозга (как впячивание дна III мозгового желудочка), и железистый (аденогипофиз), образующийся из впячивания верхней стенки глотки. В аденогипофизе выделяют три части (лопасти, доли): главную (переднюю, расположенную на периферии), переходную (наибольшую) и промежуточную (рис. 32). Аденогипофиз является центральной железой эндокринной системы. В железистой паренхиме его ролей вырабатывается секрет, содержащий ряд гормонов, стимулирующих рост (соматический гормон необходим для роста костей), регулирующих функции половых желез и таким образом воздействующих на половое созревание влияющих на деятельность пигментных клеток (определяют окраску тела и прежде всего появление брачного наряда) и повышающих устойчивость рыб к высокой температуре. Удаление гипофиза влечет за собой остановку роста и созревания.
· Гормоны, выделяемые нейрогипофизом, синтезируются в ядрах гипоталамуса и переносятся в нейрогипофиз, а затем попадают в пронизывающие его капилляры. Таким образом, это нейросекреторная железа. Гормоны принимают участие в осморегуляции, вызывают нерестовые реакции.
· Наиболее интенсивное развитие гипофиза приходится на период превращения личинки в малька. У половозрелых рыб активность его неравномерна в связи с биологией размножения рыб и, в частности, с характером икрометания.
· У единовременно мечущих рыб секрет в железистых клетках накапливается почти одновременно; после выведения секрета, к моменту овуляции, гипофиз опустошается и в секреторной деятельности его наступает перерыв. В яичниках к моменту нереста заканчивается развитие овоцитов, подготавливаемых к вымету в данный сезон. Овоциты вымётываются в один прием и составляют таким образом единственную генерацию.
· У порционно нерестующих рыб секрет в клетках образуется не одновременно. Вследствие этого после вывода секрета во время первого нереста остается часть клеток, в которых процесс образования коллоида не закончился. В результате он может выделяться порциями на протяжении всего нерестового периода.
· В свою очередь, овоциты, подготавливаемые к вымету в данный сезон, развиваются также асинхронно: к моменту первого нереста в яичниках содержатся не только созревшие овоциты, но и те, развитие которых ещё не завершено; такие овоциты созревают через некоторое время после выведения первой генерации овоцитов, т. е. первой порции икры. Так образуется несколько порций икры.
Схема эндокринной регуляции полового цикла у половозрелых рыб:
Окружающая среда (температура, свет, присутствие особей другого пола, наличие нерестового субстрата и т. д.) --> промежуточный мозг (гипоталамус) --> гипофиз --> гонады
· Исследования путей стимуляции созревания рыб привели почти одновременно, но независимо друг от друга, бразильских (Иеринги Кардозо) и советских ученых (Гербильский и его школа, 1932–1934) к разработке метода гипофизарных инъекций производителям для ускорения перевода их в текучее состояние.
· Возможность перевода производителей из IV в V стадию зрелости с помощью инъекций гипофиза, использование для этих целей гипофизов, полученных как от рыб тех же видов, так и других, включая и малоценных (гетерогенная инъекция), возможность применения заготовленных заранее, ацетонированных гипофизов (действие которых сходно с действием свежих) позволило в значительной мере управлять процессом созревания рыб и тем самым увеличивать размах рыбоводных работ по воспроизводству ценных видов. Гипофизарные инъекции широко применяют при искусственном разведении осетровых и многих карповых рыб (белый амур, белый и пестрый толстолобики, карп и др. ).
· Нейросекреторным органом у костистых рыб является и урогипофиз, находящийся в каудальной области спинного мозга и участвующий в осморегуляции.
· Щитовидная железа расположена в области глотки, около брюшной аорты. У одних рыб (некоторые акулы, лососевые и т. д. ) она является относительно плотным парным образованием, состоящим из фолликулов, выделяющих гормоны; у других (окуневые, карповые и некоторые другие) железистые клетки не образуют оформленного органа, а лежат диффузно в соединительной ткани.
· Секреторная деятельность щитовидной железы начинается очень рано: например, у личинок осетра на 2-й день после выклева железа, хотя и не вполне сформированная, обнаруживает активную секреторную деятельность, а на 15-й день формирование фолликулов почти заканчивается. Содержащие коллоид фолликулы обнаруживаются у 4-дневных личинок севрюги.
· В дальнейшем железа периодически выделяет скапливающийся секрет, причем усиление ее деятельности отмечается у молоди вовремя метаморфоза, а у половозрелых рыб – в преднерестовый период, до появления брачного наряда. Максимум активности совпадает с моментом овуляции.
· Активность щитовидной железы меняется в течение жизни, постепенно падая в процессе старения, а также в зависимости от обеспеченности рыб пищей: недокорм вызывает усиление функции. У самок щитовидная железа развита сильнее, чем у самцов, однако у самцов она более активна.
· Как и у высших животных, у рыб щитовидной железе принадлежит важная роль в регуляции обмена веществ: процессов роста и дифференцировки, углеводного обмена, осморегуляции, поддержании нормальной деятельности нервных центров, коры надпочечников, половых желез. Некоторые авторы отмечают ее более высокую гормональную активность. Добавление препарата щитовидной железы в корм ускоряет развитие молоди. При нарушении функции железы появляется зоб, для лечения которого применяют йодистые и другие препараты.
· Половые железы – яичники и семенники – выделяют половые гормоны. Секреция их периодична, наибольшее количество гормонов образуется в период зрелости гонад. С этими гормонами связывают появление брачного наряда.
· В яичниках акул и речного угря, а также в плазме крови акул обнаружены гормоны 17b-эстрадиол и эстерон, локализующиеся преимущественно в яйцеклетках, меньше – в ткани яичника. У самцов акул и лосося определены дезоксикортикостерон, прогестерон и т. д.
· Как и у высших позвоночных животных, у рыб существует функциональная зависимость между гипофизом, щитовидной железой и гонадами. В преднерестовый и нерестовый периоды созревание гонад направляется активностью гипофиза и щитовидной железы; деятельность этих желез, в свою очередь, взаимосвязана.
· Поджелудочная железа у костистых, как указывалось, выполняет двойную функцию – железы внешней секреции (выделение ферментов) и внутренней (выделение инсулина).
· Образование инсулина локализовано в островках Лангерганса, вкрапленных в ткань печени.
· Ультимобранхиалъные – супраперибранхиальные, или подпищеводные, железы – обнаружены у многих рыб, как морских, так и пресноводных. Это – парные или непарные образования, лежащие, например, у щук и лососевых по бокам пищевода. Клетки железы секретируют гормон кальцитонин, который препятствует резорбции из костей кальция и таким образом не дает повышаться его концентрации в крови.
· Надпочечники. В отличие от высших животных у рыб мозговое и корковое вещество анатомически разобщено и не образует единого органа. У костистых рыб оно располагается в разных участках почки. Корковое вещество (соответствующее кортикальной ткани высших позвоночных) внедрено в переднюю часть почки и носит название интерреналовой ткани. В корковом веществе обнаружены те же вещества, что и у других позвоночных, но содержание, например, липидов, фосфолипидов, холестерина, аскорбиновой кислоты у рыб выше.
· Гормоны коркового слоя оказывают многостороннее влияние на жизнедеятельность организма: глюкокортикоиды (у рыб обнаружены кортизол, кортизон, 11-дезоксикортизол), половые гормоны и другие, как предполагают, принимают участие в развитии скелета, мышц, половом поведении и т. д. Изъятие интерреналовой ткани ведет к остановке дыхания – ещё до остановки сердца. Кортизол участвует в осморегуляции.
· Мозговому веществу надпочечников высших животных у рыб соответствует хромаффинная ткань, отдельные клетки которой разбросаны в ткани почек. Выделяемый ими гормон адреналин воздействует на сосудистую и мышечную систему: увеличивает возбудимость и силу пульсации сердца, вызывает расширение и сужение сосудов. С повышением концентрации адреналина в крови связывают возникновение реакции тревоги.
Ядоносные и ядовитые рыбы
· Рыб, отличающихся ядовитыми свойствами, разделяют на ядоносных и ядовитых. Ядоносные рыбы имеют ядоносный аппарат –шипы и ядовитые железы, расположенные у основания шипов (на пример, у Myoxocephalus scorpius в период икрометания) или в желобках шипов и плавниковых лучей (Scorpaena, Frachinus, Amiurus, Sebastes и др. ). Сила действия ядов различна – от образования в месте укола нарыва до расстройства дыхания и сердечной деятельности и смерти (в тяжелых случаях поражения Trachurus). При употреблении в пищу эти рыбы безвредны. Рыбы, ткани и органы которых ядовиты по химическому составу, относятся к ядовитыми употребляться в пищу не должны. Они особенно многочисленны в тропиках. У акулы Carcharinus glaucus ядовита печень, у скалозуба Tetrodon – яичники и икра. В нашей фауне у маринки Schizothorax и османа Diptychus ядовиты икра и брюшина, у усача Barbus и храмули Varicorhynus икра оказывает слабительное действие. Яд ядовитых рыб действует на дыхательные и вазомоторные центры, не разрушается при кипячении. У некоторых рыб ядовита кровь (угри Muraena, Anguilla, Conger, а также минога, линь, тунец, карп и др. ). Ядовитые свойства проявляются при инъекции кровяной сыворотки этих рыб; они пропадают при нагревании под действием кислот и щелочей.
· Отравления несвежей рыбой связаны с появлением в ней ядовитых продуктов жизнедеятельности гнилостных бактерий. Специфический же “рыбный яд” образуется в доброкачественной рыбе (преимущественно осетровых и белорыбице) как продукт жизнедеятельности анаэробной бактерии Bacillus ichthyismi (близкой к В. botulinus). Действие яда проявляется при употреблении сырой (в том числе соленой) рыбы.
Глава 2. Основные звенья жизненного цикла рыб
Размножение и развитие рыб
· Развитие организма представляет собой совокупность количественных и качественных изменений в результате взаимодействия организма со средой. В индивидуальном развитии рыб можно выделить ряд крупных отрезков – периодов, каждый из которых характеризуется общими для разных видов свойствами.
· I. Эмбриональный период – oт момента оплодотворения яйца до перехода молоди на внешнее питание. Эмбрион питается за счёт желтка – запаса пищи, полученного от материнского организма. Этот период подразделяется на два подпериода:
· 1) подпериод икринки, или собственно эмбриона, когда развитие происходит в оболочке;
· 2) подпериод свободного эмбриона (предличинки), когда развитие идет вне оболочки.
· II. Личиночный период начинается с момента перехода на питание внешней пищей; внешний облик и внутреннее строение ещё не приняли формы взрослого организма. У личинок имеются специфические личиночные органы, которые в дальнейшем пропадают.
· III. Мальковый период – внешний облик близок к облику взрослого организма. Исчезают личиночные органы, появляются характерные для взрослых органы и функции. Половые органы почти неразвиты. Энергетические ресурсы расходуются главным образом нарост. Вторичнополовые признаки обычно отсутствуют.
· IV. Период полувзрослого (неполовозрелого) организма: начинается более или менее быстрое развитие половых желез и вторично-половых признаков, но организм ещё не способен к размножению.
· V. Период взрослого (половозрелого организма – состояние, при котором в определённый период года организм способен воспроизводить себе подобных; вторичнополовые признаки, если они свойственны данному виду, имеются. Энергия тратится преимущественно. На развитие половой системы и создание запасов для поддержания жизнедеятельности во время миграций, зимовок, размножения.
· VI. Период старости – половая функция затухает; рост в длину прекращается или крайне замедляется.
· В пределах подпериода или периода выделяют этапы. Теория этапности развития рыб разработана отечественными учеными школы академика А. Н. Северцова С. Г. Крыжановским и В. В. Васнецовым. На каждом этапе организм характеризуется специфическими приспособлениями к среде, т. е. определёнными особенностями строения, дыхания, питания, роста. На протяжении этапа организм растет, но значительных изменений в его строении и отношениях со средой не происходит. При этом вырабатываются свойства, обеспечивающие переход к следующему этапу. Стадия обозначает любой данный момент развития.
· Размножение – важнейший жизненный процесс, обеспечивающий существование вида. В органическом мире размножение может происходить двумя способами – бесполым и половым.
· Рыбам свойственно половое размножение, хотя у многих видов сельдей, осетровых, лососевых, карповых и некоторых других зрелые половые клетки, попав в воду, начинают развиваться партеногенетически, т. е. без оплодотворения. При этом, как правило, развитие доходит только до стадии дробления и лишь в исключительных случаях были получены жизнеспособные личинки, прожившие до полного рассасывания желточного мешка (салака, сахалинская сельдь, налим, чебачок, окунь).
· Некоторым видам рыб (серебряный карась Carassius auratusgibelio, молинизия Molinisia formosa) свойствен совершенно необычный способ размножения – гиногенез. У этих видов во многих районах ареала популяции состоят только из самок (самцы отсутствуют или единичны и неполноценны в половом отношении). В таких однополых популяциях нерест самок проходит при участии самцов других видов. При этом виде размножения проникновение спермия в яйцеклетку является необходимым условием развития. Однако слияния ядер спермия и яйцеклетки не происходит и ядро яйцеклетки становится ядром зиготы (ядро спермия генетически инактивируется). В результате в потомстве появляются только самки без внешних признаков тех самцов, которые принимали участие в нересте. Цитогенетической основой этого процесса является триплоидия самок из однополых популяций (при специфическом ходе делений созревания).
· Размножение и развитие рыб отличаются рядом специфических особенностей, обусловленных водным образом жизни.
· У большинства рыб осеменение наружное. В отличие от наземных животных зрелые половые клетки рыб выводятся в воду, здесь происходит оплодотворение икры и дальнейшее ее развитие. Осеменение, оплодотворение и инкубация икры в воде, вне материнского организма, влечет за собой большую гибель потомства на ранних стадиях развития. Для обеспечения сохранения вида в процессе эволюции у рыб выработалась или большая плодовитость, или забота о потомстве.
· Плододовитость рыб много выше, чем у наземных позвоночных. Это приспособительное свойство вида к условиям существования. Количество икры, откладываемой разными видами, очень сильно варьирует – от нескольких штук у полярной акулы до 200 млн. у морской щуки и 300 млн. у луны-рыбы. Наиболее плодовиты рыбы, откладывающие плавающую пелагическую икру; затем следуют рыбы, икра которых развивается приклеенной к растениям. У рыб, прячущих или охраняющих свою икру, плодовитость невелика.
· Наблюдается обратная зависимость между индивидуальной плодовитостью и размерами икринок: у рыб с крупной икрой она ниже, с мелкой – выше (у кеты диаметр икринок 7–8 мм, плодовитость2–4 тыс. шт. , у трески диаметр икринок 1,1 –1,7 мм, плодовитость до 10 млн. шт. ).
· Сильнейшее влияние на плодовитость оказывает обеспеченность рыб пищей. У рыб одного размера плодовитость значительно выше в благоприятных условиях питания – в кормные годы, в разреженных популяциях. Кроме того, у одного и того же вида плодовитость зависит от размера и возраста рыбы. У одной и той же особи плодовитость –при прочих равных условиях – по мере роста сначала увеличивается, затем к старости уменьшается, несмотря на продолжающийся абсолютный рост.
· Различают плодовитость индивидуальную, относительную и рабочую. Индивидуальная, или абсолютная, или общая, плодовитость –это общее количество икры, выметываемое самкой за один нерестовый период при благоприятных условиях. Например, у 6-летнегокарпа она составляет в среднем около 900 тыс. шт. Относительная плодовитость – количество икры, приходящееся на единицу массы тела самки; у карпа 180 тыс. шт/кг массы тела; этот показатель особенно удобен для сравнения, он четко показывает, как изменяется количество икры с ростом рыбы: до определённого возраста оно возрастает, потом снижается. Рабочая плодовитость – количество икры, получаемое от одной самки при проведении искусственного осеменения в рыбоводной практике. Для определения плодовитости берут пробу икры в стадии ее наибольшего развития, т. е. незадолго до нереста.
· Наступление половой зрелости у разных видов происходит в разном возрасте, причем во многих случаях самцы созревают на год раньше самок. Наиболее скороспелые промысловые рыбы, с коротким жизненным циклом (некоторые бычки, каспийская килька, хамса, снеток), созревают в годовалом возрасте. Рыбы с продолжительным жизненным циклом, например осетровые, становятся половозрелыми в 7–8 (севрюга), 12–13 (осетр) и даже 18–20 лет (белуга и калуга).
· У рыб одного и того же вида половозрелость может наступать в разном возрасте, в зависимости прежде всего от температуры выращивания и наличия пищи. Карп в средней полосе созревает в возрасте 4–5 лет, в южных районах – в 2 года и затем нерестует1 раз в год, в тропиках – на Кубе – в возрасте 6–9 месяцев и нерестует с полугодовым интервалом. У рыб одного вида ритм полового созревания в тропиках иной, чем в средней полосе (рис. 33). Обычно время наступления половой зрелости связано с достижением особью определённых размеров. Чем медленнее рыба растет, тем позднее созревает. Соотношение полов у разных видов изменчиво, но у большинства близко к 1: 1, кроме тех, у которых наблюдается гиногенез.
· Вторичных половых признаков у большинства рыб нет, поэтому у них самки и самцы внешне не различаются. Однако половой диморфизм проявляется в различной величине самок и самцов: среди одновозрастных рыб самки бывают крупнее самцов, за исключением некоторых видов – мойвы, нерки, канального сома. Однако у многих рыб половой диморфизм становится Заметным в преднерестовый период, при созревании, когда появляется так называемый брачный наряд – в подавляющем большинстве случаев у самцов. У карповых и сигов на голове и теле образуется жемчужная сыпь, у хариусов краснеют плавники, у гольцов на теле появляются яркие пятна, у лососей изменяются челюсти и возникает горб (нерка, горбуша), появляются лиловые пятна вдоль тела (кета) и т. д. После нереста брачный наряд пропадает, однако, например, у дальневосточных лососей, угрей, сельди-черноспинки изменения в организме настолько сильны и необратимы, что после первого нереста рыбы погибают. У некоторых видов нашей фауны вторичные половые признаки выражены достаточно четко.
· Они проявляются в большинстве случаев в величине плавников: у некоторых сомов, пескаря Gobio gobio, байкальской желтокрылки Cottocomephorus grewingkii, грудные плавники у самцов больше, чем у самок; у самцов линей Tinca tinca брюшные плавники больше и вторые лучи их толще; у самцов хариуса Thymallus thymallus и четырехрогого подкаменщика Myoxocephalus quadricornis длиннее спинные плавники. У полярных камбал рода Liopsetta самцы имеют ктеноидную, а самки – циклоидную чешую. У некоторых глубоководных рыб (Ceratiidae) мелкие самцы прирастают к телу самки.
· Очень своеобразны преднерестовые изменения у горчаков Rhodeinae, откладывающих икру в мантийную полость двустворчатых моллюсков: у самцов окраска становится очень яркой, а у самок вырастает длинный яйцеклад.
· По срокам икрометания рыб нашей фауны разделяют на:
· а) весенне-нерестующих (сельди, радужная форель, щука, окунь, плотва, орфа);
· б) летне-нерестующих (сазан, карп, линь, краснопёрка);
· в) осенне-зимне-нерестующих (многие лососи, сиги, налим, навага).
· Это деление в известной мере условно–один и тот же вид в разных районах нерестует в разное время: карп нерестует в средней полосе в мае–июне, на островах Ява и Куба – круглый год.
· Время нереста сильно варьирует в течение суток: лососи, налим, хамса обычно вымётывают икру ночью, анчоус – вечером, карп нерестует чаще всего на зорях.
· Изменения половых желез рыб в течение года (годичные половые циклы) проходят по одной схеме. Однако у разных видов имеются особенности в созревании и прежде всего в продолжительности разных стадий зрелости.
· По продолжительности периода икрометания выделяют две группы рыб: с единовременным и порционным нерестом. У рыб единовременного икрометания икра откладывается сразу, единовременно: в короткий срок (одно утро) нерестуют вобла, окунь. Многие тропические рыбки выметывают икру в течение часа. Вся икра таких рыб, предназначенная к вымету в данный сезон, созревает сразу и полностью выметывается.
· Другие рыбы откладывают икру в несколько приемов, отдельными порциями, с промежутками в 7–10 дней. Типичный представитель – каспийские сельди. У них в гонадах икра созревает и выметывается последовательными порциями. В результате увеличивается индивидуальная плодовитость: при порционном икрометании за сезон самка выметывает в 2–3 раза больше икры, чем при единовременном.
· Порционность икрометания характерна главным образом для рыб тропиков и субтропиков, в умеренных широтах их меньше, в Арктике – почти нет.
· Существуют рыбы, которые хотя и не имеют резко выраженного порционного икрометания, но нерестовый период их (одной особи) растягивается на несколько дней, т. е. икра также выметывается в несколько приемов (лещ, иногда карп). Некоторые рыбы в южной части своего ареала нерестуют порционно, в северной – единовременно (лещ, карп).
· Порционное икрометание способствует увеличению плодовитости рыб и обеспеченности потомства пищей, а также лучшей выживаемости молоди в неблагоприятных условиях обитания. Например, в водоемах с колеблющимся уровнем значительно больше видов рыб с порционным нерестом.
· Выметанные икринки у подавляющего большинства рыб шаровидны, но есть и овальные (хамса), сигаровидные (бычки, ротан) и даже каплевидные и цилиндрические (некоторые бычки). Окраска икринок у большинства видов желтоватая, оранжевая разных оттенков, у осетровых – черная, у бычков – зеленая. Желтоватый и оранжевый цвет обусловлен присутствием каротиноидов. Размеры икринок сильно варьируют: у некоторых сельдей, камбал икринки имеют менее 1 мм в диаметре, у акул – до 8–9 см и выше, причем они увеличиваются по мере продвижения вида к северу и на глубины.
Величина икринок, мм
· Плащеносная акула Chlamydoselachus anguineus 90 – 97
· Ручьевая форель Salmo trutta morpha fario 4,0 – 6,5
· Черноморский шпрот Sprattus sprattus 0,9 – 1,15
· Сельдь-черноспинка Caspialosa kessleri 2,87 – 3,93
· Карп Cyprinus carpio 0,9 – 1,5
· Линь Tinca tinca 1,0 – 1,2
· Чехонь Pelecus cultratus 3,8 – 5,9
· Речная камбала Pleuronectes flesus 0,78 – 1,3
· Окунь Perca fluviatilis 2,0 – 2,5
· Судак Lucioperca lucioperca 1,2 – 1,4
· Щука Esox lucius 2,5 – 3,0
· Икринки, выметанные и развивающиеся в разных экологических условиях, обладают рядом особенностей, которые способствуют их приспособленности к среде, (рис. 34). В толще воды развиваются плавающие, или пелагические, икринки, на дне или на субстрате –донные, или демерсальные.
· У пелагических икринок, развивающихся в толще воды, увеличение плавучести обеспечивается рядом приспособлений. К ним относятся: оводнение желтка (в морских пелагических икринках содержание воды доходит до 85–97%, благодаря чему они легче морской воды, тогда как в донных – до 60–76%), увеличение перивителлинового пространства за счет наличия в желтке жировых капель (многие сельди, камбалы) или образование выростов, облегчающих удерживание икринки в толще воды (сайра и др. ).
· У чехони, дальневосточных растительноядных рыб, проходных сельдей икринки полупелагические; они развиваются в толще воды, на течении, в реке, но в стоячей воде тонут.
· Икринки, откладываемые на субстрат (вегетирующие или отмершие растения, камни, коряги и т. д. ), часто обладают клейкими оболочками (осетровые, атлантическая и тихоокеанская сельди, карп, карась, рыбец и т. д. ) или снабжены нитевидными или крючковидными отростками, которыми они прикрепляются к субстрату. Икринки часто откладываются компактно, и кладки имеют характерную форму. Например, у окуня икринки окружены вязким студенистым веществом, кладки имеют вид длинных (2–3 м) лент (рис. 35). Однако они могут и не прикрепляться к субстрату (лососевые, налим). Донные икринки свойственны подавляющему большинству пресноводных рыб или морским, нерестующим в прибрежной зоне. Количество желтка и плазмы в икринках разных видов рыб не одинаково. По их соотношению яйца костистых рыб делят на олигоплазматические (содержащие мало плазмы и много желтка) и полиплазматические (богатые плазмой и бедные желтком).
· Резервный материал для питания зародыша – желток овоцита –состоит преимущественно из белков, основная масса которых представлена липофосфопротеидами (ихтулин) и небольшим количеством альбумина, и липидов (главным образом фосфатидов, прежде всего лецитина, а также холестерина); имеется небольшое количество полисахаридов и нейтральных жиров.
· У многих рыб цитоплазма овоцита содержит жировые капли, состоящие преимущественно из нейтральных жиров – глицеридов. Яйца рыб характеризуются большим количеством воды. Сильно колеблется в них содержание белков (от 12–14 до 29–30% от сырой массы) и жиров (от 1–2 до 22% сырой массы). При этом количество белков преобладает над количеством жиров (отношение белки/жиры, например у пеляди составляет 1,17, у форели – 3,25, у сазана – 4,15, а у щуки и судака – 21,19–21,66).
· Различна и калорийность икры, например у осетра и лосося 25522–25941 Дж/г, у барабули 16 318 Дж/г сухого вещества. Содержание углеводов в яйцах рыб незначительно: глыбки гликогена обнаружены (цитохимически) у осетровых, лосося, пеляди, карпа.
· Таким образом, главным источником энергии при развитии зародыша являются белки, за счет которых покрывается до 70% расходуемой энергии. Жиры, в отличие от жиров яиц птиц, расходуются в меньшей степени. При этом в пелагических икринках запас энергетических веществ меньше, в донных – больше.
· Зрелый сперматозоид представляет собой клетку с малым количеством плазмы. В нем различают головку, среднюю часть и хвост (рис. 36). Форма головки различна: в виде шара, яйца, желудя (у большинства костистых рыб), палочки (у осетровых и некоторых костистых), копья (у двоякодышащих), цилиндра (у акуловых, кистепёрых). В головке помещается ядро. Впереди ядра у акулообразных, осетровых и некоторых других рыб располагается акросома; у костистых акросомы нет. Ядерная часть головки сперматозоида состоит преимущественно из дезоксирибонуклеопротеида (нейтральная соль ДНК с основным белком – протамином) и небольшого количества РНК. Концентрация ДНК в головке (ядре) составляет 38,1% (карп), 48,4% (семга) и отражает количество ДНК в гаплоидном наборе хромосом. Протамины состоят из 6–8 аминокислот, среди которых преобладает аргинин. В средней части сперматозоидов обнаружены митохондрии, играющие основную роль в снабжении клетки энергией. В хвостовой части выявлены белки, лецитин, жиры и холестерин. Сперматозоиды большинства костистых рыб имеют общую длину 40–60 мкм (головка 2–3мкм).
· Сперма, выделяемая самцом, состоит из сперматозоидов, погруженных в спермиальную жидкость, сходную по составу с физиологическим раствором. В момент выхода из организма сперматозоиды ещё неподвижны, обмен их понижен.
· У одного и того же самца сперматозоиды качественно неодинаковы. Прежде всего они различаются по величине: в эякуляте при помощи центрифугирования можно выделить три группы сперматозоидов – мелких (легких), крупных (тяжелых), промежуточных (средних). Они различаются и по биологическим свойствам, в частности по характеру гамет: среди крупных сперматозоидов в большом количестве встречаются X-гаметы, среди мелких – Y-гаметы. Вследствие этого из икры, оплодотворенной крупными сперматозоидами, рождаются преимущественно самки, а мелкими – самцы.
· Эти данные используют для разработки направленного формирования пола у рыб, которое имеет важное значение в рыбоводстве. Количество спермы, которое единовременно выделяет самец при нересте, и концентрация эякулятов различны у разных видов (табл. 6). Оно зависит от комплекса внутренних и внешних факторов: размеров, возраста и состояния самцов, условий нереста – течений, температуры, соотношения самок и самцов на нерестилищах и т. д.
· В спермиальной жидкости сперматозоиды неподвижны. При соприкосновении с водой их обмен резко возрастает, поглощение кислорода увеличивается больше чем вдвое, содержание АТФ – больше чем втрое; сперматозоиды активируются и начинают бурно двигаться со скоростью, наблюдаемой у сперматозоидов млекопитающих (164–330 мкм/с). Встретив икринки, они проникают в них через микропиле, после чего происходит оплодотворение. Однако в воде сперматозоиды недолговечны. Энергетические ресурсы их иссякают, первоначальное поступательное движение замедляется, становится колебательным, затем прекращается и они погибают. Продолжительность подвижности сперматозоидов колеблется в зависимости от солености среды, в которой проходит нерест, и температуры; в соленой воде она значительно дольше: до нескольких суток у тихоокеанской сельди Clupea harengus pallasi, в пресной воде у большинства рыб – карповых, лососевых, окуневых – не больше 1–3 мин.
Таблица 6 Характеристика спермы разных видов рыб (по Казакову, 1978, с сокращениями) .
| Вид рыбы | Единовремен ный объём спермы, см3 | Концентрация сперматозоидов, млн. /мм |
Продолжительность активности сперматозоидов, с
|
||
| Радужная форель Пелядь Карп Белый амур | 1,0-23,0 0,2-3,2 2,9-12,5 20-30 | 20,4 7,6 16,5 33,1 |
|
· Между подвижностью сперматозоидов и их оплодотворяющей способностью наблюдается прямая зависимость: уменьшение скорости движения сопровождается падением процента оплодотворённости икры. При этом имеет значение поступательное движение (колебательное движение не дает возможности сперматозоиду проникнуть в икринку).
· В эякуляте без добавления воды неподвижные сперматозоиды долго сохраняют оплодотворяющую способность – до нескольких часов и даже суток. На этом основаны хранение и перевозка “сухой” спермы.
· Рыбы почти всех видов раздельнополы. Органический гермафродитизм свойствен миксинам. Среди костистых рыб обычно являются гермафродитами лишь морские окуни Serranus и морские караси Sparidae. Изредка гермафродиты появляются во многих семействах как среди морских, так и пресноводных форм (у сельдевых, лососевых, щуковых, карповых, окуневых и др. ). При этом, например, у кеты и кефали в гонадах чередуются участки яичников и семенников. Крайне редки сообщения о гермафродитизме карпа. В одном из таких случаев описано выделение гермафродитом одновременно икры и спермы. При этом самооплодотворение сопровождалось значительным отходом икры (развилось 29% зародышей), тогда как при осеменении спермой гермафродита икры другой самки развивалось 98% икринок.
· У рыб может происходить изменение, превращение (реверсия) пола. Например, молодь радужной форели на ранних стадиях (в возрасте 135–160 дней), имевшая в гонадах массу женских половых клеток, в дальнейшем развивалась в самцов. У большинства пресноводных рыб половые железы во время закладки индифферентны в отношении половой принадлежности, они как бы потенциально двуполы. Пол такой интерсексуальной особи определяется при дальнейшем развитии.
· Но превращение пола может наблюдаться и у взрослых особей. Известны случаи, когда у зубастых карпов Cyprinodontoidae половозрелые, уже ранее нерестовавшие самки вдруг превращались в самцов и становились способными оплодотворять икру; у некоторых рыб в течение жизни перестройка пола наблюдается неоднократно.
· У рыб имеет место избирательность оплодотворения. Поэтому использование при осеменении икры спермы двух (или более) особей повышает оплодотворяемость икры.
· Рыбы размножаются в самых различных условиях. В связи с особенностями, строения, размножения на различном нерестовом субстрате и развития рыб выделяют следующие экологические группы: Литофилы – размножаются на каменистом грунте, обычно в реках на течении или на дне олиготрофных озер или прибрежных участков морей, обычно в благоприятных условиях кислородного режима. Это осетры, лососи, подусты и др.
· Фитофилы – размножаются среди растительности, откладывая икру в стоячей или слаботекучей воде на отмершие или вегетирующие растения. При этом кислородные условия сильно варьируют. К этой группе принадлежат щука, сазан, лещ, плотва, окунь и др.
· Псаммофилы – откладывают икру на песок, иногда прикрепляя ее к корешкам растений. Часто оболочки икринок инкрустируются песком. Развиваются обычно в благоприятных условиях дыхания. К этой группе принадлежат пескари, некоторые гольцы и др.
· Пелагофилы – выметывают икру в толщу воды. Икра и свободные эмбрионы развиваются, свободно плавая в толще воды, обычно в благоприятных для дыхания условиях. В эту группу входят почти все виды сельдей, тресковых, камбал, некоторые карповые (чехонь, толстолобики, амуры и др. ).
· Остракофилы – откладывают икру внутрь мантийной полость моллюсков и иногда под панцири крабов и других животных. Икра обычно развивается в не особенно благоприятных условиях дыхания. Это некоторые пескари, горчаки и др.
· Эта классификация охватывает не всех рыб, имеются промежуточные формы: рыбец может нерестовать и на растительности, и на камнях, т. е. как фитофильная и литофильная рыба.
· В зависимости от кислородного режима нерестилищ, при котором обычно инкубируется икра, развиваются “дыхательные” пигменты – каротиноиды желтоватого и красноватого цвета. Чем хуже условия дыхания, тем сильнее пигментирована икра. Поэтому самая яркая окраска присуща икре фитофильных и литофильных рыб, а самая светлая – икре пелагофилов.
· Большинство рыб не заботится о потомстве. Нередки случаи, когда родители даже поедают собственную икру и особенно молодь.
· Каннибализм встречается у гамбузии, наваги, даже карпа. Поэтому целесообразно в целях сохранения молоди вылавливать производителей из нерестовых прудов. Однако немало видов рыб, заботящихся о потомстве. При этом охрана потомства в большинстве случаев выпадает на долю самцов.
· Примеры заботы о потомстве интересны и разнообразны: колюшка (самец) строит гнездо из кусочков травинок, склеиваемых выделениями почек. Гнездо имеет сначала два отверстия, а после наполнения его икрой (несколькими самками) самец закрывает одно отверстие и остается охранять его, аэрируя воду движениями плавников. После выклева молоди самец в течение нескольких дней следит за тем, чтобы она находилась в гнезде и возвращает туда выплывающих, захватывая их ртом (рис. 37). Тиляпии вынашивают икру во рту (самки) и некоторое время после выклева забирают молодь в рот при опасности. У морской иглы и морского конька икра инкубируется в складке или сумке на брюшке самцов. Лабиринтовые рыбки строят гнездо из пены – пузырьков воздуха и слюнообразного секрета. Хотя молодь в гнезде появляется через сутки, самец охраняет его до тех пор, пока рыбки окончательно не оформятся. Вообще постройка гнезд разной сложности встречается у рыб нередко. Форель и лосось выкапывают в грунте несколько ямок, а отложенную икру засыпают песком и гравием движениями хвоста (устраивая так называемые нерестовые бугры). Некоторые бычки, сомы устраивают гнезда из камешков и кусочков растений; пинагор охраняет комок икры, отложенной у полосы прибоя, и вовремя отлива поливает его водой из рта. Судак строит гнездо из кусочков корней или расчищая каменистый участок; он кусает протянутую к гнезду руку, и отогнать его не удается; движением грудных плавников он создает ток воды, смывающий ил с икринок.
· Наиболее совершенной формой заботы о потомстве является живорождение. При этом плодовитость обычно бывает мала –несколько десятков особей. По сути это – яйцеживорождение с задержкой потомства в половых путях самки, до рассасывания желточного мешка. Оно присуще многим акуловым, а среди костисты хрыб – бельдюге Zoarces viviparus, морскому окуню Sebastes marinus, гамбузии Gambusia affinis, гуппи Lebistes reticulatus, меченосцу Xinophorus hellery, из карповых – усачу Barbus viviparus.
Эмбриональное и раннее постэмбриональное развитие рыб
· В первые мгновения после оплодотворения оболочки икринки прилегают к поверхности желтка. Затем кортикальные альвеолы, располагающиеся в поверхностном слое цитоплазмы, лопаются, их содержимое выделяется под оболочку и она отслаивается от желтка. Начинается оводнение (набухание) икринки, в процессе которого между желтком и оболочкой образуется перивителлиновое пространство, заполненное жидкостью. Эта жидкость обеспечивает обмен зародыша и защищает его от воздействия внешней среды.
· Перивителлиновое пространство образуется и в неоплодотворенной икринке после попадания ее в воду. Так как перивителлиновое пространство препятствует проникновению сперматозоидов, то после его образования икринка теряет способность к оплодотворению.
· Внешняя оболочка икринки многих рыб выделяет клейкое вещество, благодаря которому в естественных условиях икринки прилипают к субстрату. После набухания прочность оболочек возрастает. Яйца костистых рыб относятся к телолецитальному типу. В них желток распределен неравномерно: ядро и плазма располагаются на анимальном полюсе, а желток концентрируется в противоположной части клетки на вегетативном полюсе. В результате дробление охватывает не всю клетку, а только бластодиск (неполное, или дискоидальное, дробление, при котором борозды дробления проходят только по бластодиску).
· Внешним признаком развития икринки является скопление плазмы на анимальном полюсе и образование бластодиска. Развитие идёт по общеизвестной схеме: дробление бластодиска (с образованием сначала крупноклеточной, затем мелкоклеточной морулы); появление бластулы, внутри которой имеется первичная полость тела – бластоцель; в результате продолжающегося размножения клеток – наступление гаструляции, в процессе которой клетки анимального полюса надвигаются на желток (обрастание желтка), образуется два зародышевых листка (экто и энтодерма); полость гаструлы представляет собой первичную полость кишечника. Затем между двумя эмбриональными пластами образуется третий (мезодерма); внутри мезодермы развивается вторичная полость тела, или целом. Далее зародышевые листки дифференцируются на зачатки тканей и органов: из эктодермы формируются покровы (эпидермис), нервная система; из энтодермы – кишечник и связанные с ним органы; из мезодермы –внутренний скелет, мускулатура, соединительнотканный слой кожи, аорта и кардинальные вены, эндокардий сердца и др.
· Эмбриональный период развития рыб не заканчивается выходом зародыша из оболочки. Он продолжается в течение некоторого времени после выклева, пока предличинка, или свободный эмбрион, обладая ещё рядом эмбриональных особенностей строения органов дыхания, кровообращения, пищеварения и других систем, проходит заключительные этапы эмбрионального развития. После того как начинают функционировать жаберная, пищеварительная и другие системы, деятельность эмбриональных органов прекращается и соответственно кончается период эмбрионального развития.
· Следующий период – личиночный – начинается с момента перехода молоди на активное питание внешней пищей. Сначала питание смешанное – остатками желточного мешка и частично внешней пищей, затем полностью экзогенное. Имеются временные личиночные органы (непарная плавниковая кайма, наружные жабры и т. д.), отсутствуют многие органы взрослой рыбы.
· При переходе в следующий период развития – мальковый молодь приобретает форму взрослой рыбы; появляется чешуя, характерные для взрослого органы и функции (например, брюшные плавники и жаберное дыхание через рот), но некоторые органы могут ещё отсутствовать, например каналы боковой линии. Личиночные органы исчезают.
· Для примера рассмотрим развитие карпа в нерестовом пруду (при температуре воды 20–22°С рис. 38).
· В течение первых суток проходят этапы, предшествующие оформлению тела зародыша.
· Образование бластодиска (1-й этап). Начинается сразу после оплодотворения. Примерно через 30 мин в икринках между желтком и наружной оболочкой возникает перивителлиновое пространство, занимающее 3,4–15,4 % диаметра икринки. На анимальном полюсе икринки формируется бластодиск в виде возвышающегося над желтком светлого бугорка.
· Дробление бластодиска (2-й этап). Бластодиск разделяется бороздами дробления на бластомеры.
· Сначала наблюдается морула крупных клеток, но по мере того, как возрастает число бластомеров, размеры их уменьшаются. Примерно через 5 ч после оплодотворения наблюдается морула мелких клеток.
· Бластула (3-й этап). Бластомеры уплотняются и отодвигаются к периферии. Образуется бластула, внутри которой имеется полость – бластоцель; желток образует впячивание навстречу накрывающей его бластодерме.
· Гаструла (4-й этап). При дальнейшем размножении клеток анимального полюса происходит обрастание желтка: бластомеры как бы сползают в сторону вегетативного полюса, постепенно накрывая его; образуется зародышевый узелок; формируются зародышевые пласты, а из них зачатки органов.
· К концу первого дня после оплодотворения в икринке имеется зародыш в виде прозрачной зародышевой полоски, лежащей на желтке. Произошла закладка головного и туловищного зачатков, причём головной конец заметен резче, хвостовой конец утончается постепенно, ограничиваясь едва заметно; выявляются участки эмбрионального материала, которые дадут начало хорде, миотомам, кишечной энтодерме, нервной и другим системам.
· В течение вторых суток проходят следующие три этапа.
· Органогенез (5-й этап). Зародыш увеличивается в размерах: тело утолщается, хвостовой отдел оканчивается перед головным, немного не доходя до него. Формируются головной, туловищный, хвостовой отделы тела и основные органы и системы органов: нервная, мышечная, кишечник и т. д. Примерно через 28 ч после оплодотворения в головном отделе хорошо виден мозг, причем заметно разделение его на передний и задний отделы, четко различимы слуховые пузырьки, глаза продолговатой формы, ещё не имеющие пигмента. В туловищном отделе происходит сегментация хорды. Примерно через 32 ч. после оплодотворения хорошо заметна плавниковая кайма, начинающаяся на спинной стороне тела в задней его трети. Кайма огибает хвостовой отдел и подходит к желтку. Видны также плавниковые складочки на желтке.
· Появляется нервно-мышечная моторика (6-й этап). Зародыш начинает временами подергиваться, а затем периодически поворачивается в оболочке. Так как зародыш в это время дышит поверхностью тела (специальных органов дыхания нет), то перемешивание перивителлиновой жидкости при таких поворотах способствует улучшению газового обмена.
· Зародыш настолько увеличивается, что хвостовой отдел начинает заворачиваться по поверхности желтка, образуя спираль. В головном отделе просматриваются обонятельные ямки, глазные бокалы, хрусталики, отолиты. В глазах появляется точечный меланин. Сердечная трубка сокращается, но форменных элементов крови ещё нет. Хорошо видна кишечная трубка. Продолжается сегментация тела (в хвостовом отделе). Желточный мешок становится грушевидным.
· Начинает функционировать эмбриональная дыхательная система (7-й этап). Так как дефинитивные органы дыхания ещё не сформированы, то дыхательную функцию выполняет сеть кровеносных сосудов: Кювьеровы протоки (лежащие на передней части желточного мешка), нижняя хвостовая вена (в хвостовом отделе тела), сеть сегментальных сосудов в плавниковой кайме (в анальной ее части). В токе плазмы крови появляются форменные элементы. Заканчивается сегментация тела. Появляются грудные плавнички. Усиливается пигментация глаз. Примерно через 52 ч после оплодотворения появляются пигментные клетки над кишечной трубкой, вскоре покрывающие головку зародыша, спинной и хвостовой отделы и желточный мешок. Пигментные клетки (меланофоры) крупные, лежат близко друг к другу (группами). На голове видны зачатки жаберных крышек. На голове и желтке появляются железки вылупления.
· К концу вторых – началу третьих суток после оплодотворения начинается последний – 8-й этап развития зародыша в оболочке. Увеличиваются все части тела и просвечивающие сквозь прозрачные покровы органы. Головка зародыша частично обособляется от желтка. В слуховых пузырьках видны полукружные каналы. Отчетливо видна ротовая ямка (рот неподвижный, открытый). Оформляется жаберно-челюстной аппарат. В передней части головы видны клетки, образующие железки приклеивания. Основания грудных плавников расположены наклонно по отношению к оси тела. В плавниковой складке обособляются спинной, хвостовой и анальный участки. Усиливается пигментация тела.
· Примерно через 78 ч после оплодотворения начинается массовый выклев молоди. Выклюнувшиеся зародыши, или предличинки (этап развития А, или последний зародышевый), имеют около 5,0–5,2 мм длины (рис. 39). Обращает на себя внимание большой желточный мешок грушевидной формы и прямая (не изогнутая) хорда. Голова немного пригнута вниз. В передней части ее, ближе к глазам, имеются углубления – обонятельные ямки. Хорошо видны сегменты (их насчитывается 38), не одинаковые по величине, они постепенно уменьшаются к заднему концу тела. По спине зародыша, начиная с 9-го сегмента, тянется вдоль тела плавниковая кайма, переходящая на хвост, далее на брюшную сторону и оканчивающаяся на желточном мешке. В хвостовой части плавниковая кайма разделяется задним концом хорды на две равные половины. Плавниковая кайма узкая, недифференцированная, без выемок, расширяется только в хвостовой части, прозрачная, чуть-чуть уплотненная с прилегающей к телу стороны; в спинной и анальной частях пронизана кровеносными сосудами. Грудные плавнички подвижны. Глаза сильно пигментированы. По телу разбросаны пигментные клетки; больше всего их на голове и вдоль спинного и брюшного краев тела, лежат они и на желточном мешке. На голове и спине имеется также желтоватый пигмент.
· На переднем краю головы зародыши имеют железу приклеивания, позволяющую им прикрепляться к подводным растениям. Сквозь прозрачное тело просвечивают внутренние органы: сердце в околосердечной сумке, кишка, прямая, ещё без просвета, не вполне сформированный жаберный аппарат – только начальные жаберные дужки прикрыты намечающейся жаберной крышкой, два отолита в слуховой капсуле. Рот открыт, имеет форму ямки.
· В течение 1-х суток жизни после выклева зародыши движутся периодически; время от времени, приклеившись к растениям, они висят неподвижно, покойно; затем, оторвавшись от субстрата, проделывают несколько червеобразных движений, после чего опять приклеиваются. Таким образом чередуются состояния движения и покоя.
· При указанных температурах преобразования зародышей протекают быстро. Уже к концу первого дня их жизни (длина 6 мм) желточный мешок оказывается сильно втянутым. На 2-е сутки жизни (длина 5,9–6,7 мм) зародыши имеют сравнительно небольшой желточный мешок. Уменьшение желточного мешка происходит по всей площади соединения его с зародышем, но быстрее в передней расширенной части. В плавниковой кайме, особенно в нижней части хвостового отдела, уплотнённые участки (скопления мезенхимных клеток) становятся более значительными. Зародыши больше не приклеиваются к растениям, они постоянно плавают.
· На 3-й сутки жизни (этап развития В, или первый личиночный) при длине тела 6,2–7,8 мм у молоди остается совсем мало желтка. Хорда по-прежнему оканчивается прямо – она не изогнута. У особей длиной около 7 мм дифференциации плавниковой каймы ещё нет, но в хвостовой части в нижней половине намечаются мезенхимные тяжики. Пигментных клеток становится больше. Жаберная крышка прикрывает не все жаберные дужки. Линия основания грудных плавников становится вертикальной. Кровь начинает окрашиваться, приобретает очень слабый жёлто-розовый оттенок.
· Кишечник представляет собой едва изогнутую трубку, но уже с просветом. Молодь заглатывает воздух, плавательный пузырь (задняя камера) наполняется им и становится хорошо видным. Наполнение плавательного пузыря воздухом облегчает передвижение рыбок. Части ротового аппарата могут двигаться. Рот перемещается на конец рыла.
· Молодь переходит к активному питанию (внешней пищей). Таким образом, в это время у личинок питание смешанное: как внешней пищей, так и за счет не совсем израсходованного желточного мешка. Вследствие прозрачности тела хорошо видно содержимое кишечника.
· На 4-е сутки жизни (этап развития С1; или второй личиночный)длина личинок достигает 5,5–9,0 мм. Самые мелкие из них имеют ещё остатки желтка. Рот приобретает способность закрываться полностью.
· У личинок, достигших длины около 8,3 мм, задний конец хорды – уростиль – начинает загибаться кверху. Зачатки лучей в нижней половине хвостовой части плавниковой каймы увеличиваются. Плавниковая кайма в передней части (на спине) становится более высокой, здесь появляется сгущение мезенхимных клеток. Такое же сгущение мезенхимы наблюдается в анальной части каймы, на месте будущего анального плавника. Перед хвостом плавниковая кайма становится немного уже, тем самым намечаются границы хвоста.
· Пигментных клеток становится очень много, они крупные, разбросаны по всему телу. Особенно крупны они на спинной стороне головы. Жаберные крышки увеличиваются. Личинки уже заглатывают циклопов, босмий и других мелких ветвистоусых и веслоногих рачков.
· На 5-е сутки жизни (этап развития С2, или третий личиночный)при длине 7,0–10,1 мм личинки отличаются от предыдущих в основном тем, что у них сильнее загнут уростиль, хвост стал гетероцеркальным, в плавниковой кайме резче выделяется хвостовой отдел, в котором лучи уже сформировались; в спинном и анальном отделах плавниковой каймы сгущения мезенхимы стали плотнее. На челюстях появляются роговые зубы. В пищевом комке кроме коловраток, ветвистоусых и веслоногих рачков начинают встречаться планктонные личинки хирономид.
· На 6-е сутки жизни (длина 8,2–11,3мм) личинки своим общим видом напоминают уже больше рыбку, чем личинку. Головка из закругленной становится вытянутой. Жаберные крышки закрывают все жаберные дужки. Хвостовой отдел на плавниковой кайме ограничивается четче, мезенхимные сгущения в спинном и анальном участках каймы уплотняются. Тело личинок становится менее прозрачным, сегменты видны плохо, только в задней части.
· На 8-е сутки жизни личинки достигают длины 10–12,8 мм. Меньшие из них прошли этап развития D1, , или четвертый личиночный, у наиболее крупных развитие продвинулось до этапа D2 –пятого личиночного.
· Сегменты в теле видны совсем плохо. Уростиль сильно загибается кверху, образуя почти прямой угол с плавниковыми лучами. В плавательном пузыре обе камеры наполнены воздухом. На месте брюшных плавников появляются кожистые выросты. Плавниковая кайма ясно дифференцирована, спинной отдел ее имеет лучи, в анальном ее отделе также появляются зачатки лучей. Хорошо видны кости черепа. У наиболее крупных рыбок хвостовой отдел представляет собой сформированный хвостовой плавник (гомоцеркальный); появляется хвостовая выемка, раздваивающая плавник на верхнюю и нижнюю лопасти. Спинной плавник также вполне сформирован. Грудные и брюшные плавники ещё не имеют лучей. Все тело очень сильно пигментировано. Рот становится выдвижным. Кишечник слабо изогнут, намечается первая петля.
· На 11-е сутки жизни (длина 11,1 – 16,0 мм) у личинок спинной и анальный участки плавниковой каймы приобретают форму плавников. С хвостовым плавником они соединяются совсем узенькими перетяжками. Лопасти брюшных плавников становятся крупнее, но лучей в них ещё нет. Пигментные клетки очень крупные. В кишечнике образуется первая петля.
· На 13-е сутки жизни (длина 12–13 мм) остатки плавниковой каймы между плавниками становятся еле заметными. Тело почти непрозрачно, лишь слабо просвечивает кишечник.
· На 14-е сутки жизни (этап развития Е, или шестой личиночный)при длине тела 15–20 мм никаких следов плавниковой каймы между плавниками нет. В брюшных и грудных плавниках появились лучи. Тело непрозрачно, его почти сплошь покрывают пигментные клетки. В кишечнике стало две петли. Чешуи ещё нет. Дальнейшее развитие происходит в выростном пруду.
· В течение развития у зародыша чередуются периоды усиленного роста тканей и периоды усиленной дифференцировки их и образования новых зачатков органов. При этом меняется характер обмена веществ, в частности интенсивность водного обмена, интенсивность усвоения биогенных элементов (фосфора, кальция, углерода), аминокислотный состав тела (уменьшается число свободных аминокислот, увеличивается количество связанных), интенсивность потребления кислорода. Наиболее интенсивен обмен во время формирования органов и тканей. Чувствительность зародышей к внешним воздействиям – тряске, колебаниям температуры, содержанию кислорода – на разных стадиях развития различна. Наименее устойчивы зародыши во время усиленного формирования тканей и органов, когда обмен наиболее интенсивен. Это начало дробления, гаструляция, закрытие бластопора, начало формирования зародыша и т. д. Это обстоятельство учитывается при работах с икрой, особенно при ее перевозках.
· Инкубация икры рыб каждого вида проходит при определённых условиях внешней среды (температура, содержание кислорода и углекислоты, рН, освещенность, соленость и т. д. ).
· Исход инкубации определяется также качеством икры. Оно связано с видом рыбы, ее возрастом, условиями содержания и в предыдущий год, и особенно перед нерестом, временем взятия икры от самок, а при искусственном осеменении – с техникой проведения всех операций.
· Длительная задержка икры в полости тела самки вызывает перезревание. Перезревшая икра характеризуется пониженной оплодотворяемостью, повышенным отходом в период инкубации, увеличенным числом уродов и самцов.
· Невыметанные зрелые половые продукты рассасываются. Но это длительный процесс. Поэтому если нерест почему-либо не прошел (например, из-за похолодания), то очередной нерест на следующий год также может не состояться, так как в гонадах не успеют пройти процессы резорбции зрелых невыметанных клеток и образования клеток новых генераций.
· Продолжительность инкубации при прочих равных условиях зависит от температуры: чем она выше, тем развитие происходит быстрее.
· У рыб, выметывающих икру весной и летом, при высоких температурах, развитие длится несколько дней; у рыб с осенне-зимним нерестом – несколько месяцев.
· Успешнее всего инкубация проходит при оптимальной температуре. При повышенной она хотя и заканчивается быстрее, но молодь выклевывается мелкой и недоразвитой, а при понижении температуры зародыши более крупные, но нарушается процесс выклева. При отклонении температуры повышается количество уродов –особей с укороченным туловищем, искривлением позвоночника, водянкой околосердечной и брюшной полости, а также двухголовых экземпляров, срастающихся разными участками туловища, особей с аномалиями челюстного аппарата и т. д.
· Для учета длительности развития существует понятие “градусо-дни”. Это произведение средней температуры инкубации на число дней развития икры. Оно дает общее представление о сумме тепла, необходимого для развития молоди до выклева. Но это не постоянная величина, она имеет разные значения при разных температурах. У карпа развитие длится 54–126 градусо-дней, у радужной форели – 330–400. При неблагоприятных условиях, например при недостатке кислорода, продолжительность развития удлиняется.
Питание и упитанность рыб
· Значение питания в жизнедеятельности организма очень велико. Пища, поступающая в организм, обеспечивает на всех этапах его развития энергетические процессы, связанные с движением, ростом, созреванием, размножением. Так через потребление пищи осуществляется одна из важнейших связей организма с окружающей средой. На протяжении индивидуального развития у рыб имеют место два типа питания – эндогенное (за счет внутренних ресурсов организма) и экзогенное (за счет внешней пищи).
· Большинство рыб большую часть жизни питается экзогенно. Однако у всех рыб питание в начальный период жизни – развитие в икринке и сразу после вылупления эмбриона – происходит за счёт запасов желтка и жира в желточном мешке (эндогенное питание) . У взрослых рыб также бывают периоды эндогенного питания, например у рыб, которые не питаются зимой или живут в пересыхающих водоемах, а также у проходных рыб во время нерестовых миграций. В это время поступление пищи извне прекращается.
· Эндогенное питание поддерживает обмен веществ у рыб во время зимовки, а у мигрирующих – покрывает огромную трату энергии при их длительных передвижениях от мест нагула к местам нереста (осетровые, лососи, некоторые сельди, угри) и созревание в это время половых продуктов, т. е. в организме происходит преобразование накопленных в предыдущий период энергетических ресурсов (в первую очередь жира).
· У дальневосточных лососей и угрей этот процесс является необратимым: организм настолько истощается, что после нереста рыба погибает.
· Соотношение этих двух форм питания у разных видов различно. По разнообразию пищи среди рыб различают монофагов (потребляющих пищу одного вида), стенофагов (набор пищевых объектов невелик) и эврифагов (пища разнокачественна).
· Существует ряд классификаций рыб соответственно их питанию. Прежде всего рыб делят на мирных и хищных.
· Мирные рыбы могут питаться беспозвоночными, растительностью и детритом. Сюда относятся мирные животноядные: планктонофаги, (сельди, некоторые сиги и т. д. ) и бентософаги (лещ, некоторые сиги и др. ); фитофаги (краснопёрка, растительноядные дальневосточные карповые – толстолобик, белый амур, амурский лещ и др. ); детритофаги (закаспийская храмуля и др. )
· Хищники питаются рыбой, а при случае даже другими позвоночными. Однако это деление весьма относительно: многие рыбы всеядны (сазан, карп), иногда бентософаги могут переходить на питание планктоном, а мирные животноядные при отсутствии обычной пищи становятся хищниками.
· Приспособленность разных видов рыб к определённому виду пищи четко проявляется в строении пищеварительного тракта –рот, жаберный аппарат, глотка, кишечник. Смена пищи на протяжении онтогенеза сопровождается морфофизиологическими изменениями.
· Изменение характера питания обусловлено рядом биотических и абиотических факторов: возрастом, полом, степенью зрелости, состоянием здоровья, сезоном года и т. д.
· Возрастные особенности питания рыб. Экзогенное питание рыб начинается не сразу после выхода из икринки, а после некоторого периода желточного питания, но до того, как желток будет полностью израсходован (у карповых, окуневых и других – через несколько дней после выклева, у лососей – через несколько недель). Промежуток времени, когда молодь питается отчасти внешней пищей, а отчасти остатками желтка, называется периодом смешанного питания. Так как молодь в это время ещё очень мала, то ей доступны лишь самые мелкие формы планктона, однако уже через 1–2 дня она может захватывать и крупных его представителей. Таким образом, молодь всех рыб вначале питается зоопланктоном. Затем, по мере роста, пищевые потребности разных видов расходятся. Одни остаются планктоноядными на всю жизнь (верховка, чехонь, многие сиги, в частности пелядь), другие начинают поедать растения (фитопланктон – белый толстолобик, высшие растения – краснопёрка, белый амур и др. ), становятся бентосоядными (сазан, линь, карп и т. д. ) или рыбоядными (щука, окунь, судак и др. ); набор пищевых объектов по мере роста рыб расширяется.
· Например, вобла начинает питаться мелким фито- и зоопланктоном, затем использует зоопланктон более крупный, далее переходит к бентосным организмам – главным образом личинкам хирономид, а став взрослой, питается преимущественно моллюсками. Соответственно возрастные изменения происходят в строении пищеварительного тракта.
· Сезонные особенности питания рыб. Многообразны изменения в питании рыб в течение года. Они связаны прежде всего с температурой воды, сезонными изменениями в составе, численности и доступности пищевых организмов и их приуроченностью к определённому району.
· Например, значительно меняется в течение года роль разных пищевых организмов в питании форели (табл. 7).
Таблица 7 Сезонные изменения в составе пищи форели, % (по Строганову, 1962, с сокращениями) .
| Месяцы | I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII |
| Донные животные, всего | 98,5 | 98,3 | 97,1 | 75,8 | 53,4 | 59,1 | 70,5 | 6,4 | 26,3 | 70,3 | 98,2 | 100 |
| Из них: гаммарусы лимнеа хирономиды | 70,5 3,9 - | 41,2 23,7 0,4 | 12,1 7,7 53,8 | 8,1 6,4 2,0 | 25,3 1,6 1,6 | 4,0 0,9 - | 0,6 1,2 0,3 | - 3,7 0,5 | 10,2 3,6 0,2 | 19,4 32,7 - | 17,5 3,9 - | 29,4 - - |
| Поверхностные животные, всего | - | 0,7 | 1,7 | 21,5 | 52,5 | 32,2 | 25,0 | 93,1 | 73,4 | 17 | 1,5 | - |
· Особенности питания рыб в зависимости от мест обитания. Фауна пищевых организмов в разных водоемах не идентична, поэтому нередко характер питания одного и того же вида в них не совпадает: например, в пище сига-пыжьяна из реки Печоры преобладают моллюски, из реки Гыды — ракообразные, из реки Кары — личинки хирономид. Очень сильно разнится доля планктона, личинок хирономид и детрита в пище серебряного карася из разных водоемов.
· Половые особенности питания рыб. Отмечено различие в наборе пищевых организмов в зависимости от пола рыбы: в Азовском море у самок леща преимущественно встречались Hypaniola, у самцов — Nereis. Наиболее разительны различия у глубоководных удильщиков: самки хищничают, а паразитирующие на них самцы питаются соками тела самок.
· Величина объектов, служащих пищей рыбам, различна: от микроскопических у планктонофагов до жертвы, превышающей размеры рыбы-хищника.
· Количественные характеристики питания рыб также разнообразны. Обычно масса пищи составляет 2—25% от массы тела рыбы.
· По значению отдельных видов пищи для разных рыб условно выделяют пищу излюбленную (ограниченную 2—6 видами организмов, которые составляют 50—75% пищевого комка), заменяющую, или второстепенную (5—6 видов, образующих 15—30% содержимого пищеварительного тракта), и случайную (много видов пищевых объектов, не превышающих, однако, 4—10% от содержимого пищеварительного тракта). Существуют и другие классификации.
· Количество пищи, потребляемой ежедневно рыбой в течение жизни, неодинаково. Относительно своей массы молодь ест больше, чем взрослые и старые рыбы: у молоди севрюги масса пищи составляла 150% по сравнению с массой тела, а позднее — только 50%.
· Меняется характер питания рыб (часто очень сильно) при созревании гонад в преднерестовый период, что, видимо, имеет гормональную основу. Многие морские рыбы при приближении нереста питаются мало или совсем не питаются. Особенно резко это выражено у проходных рыб: дальневосточные лососи перестают питаться при переходе из моря в реку за 2—3 мес., а иногда за 1 год до нереста.
· Сильно ослабляется питание при заболеваниях рыб. Рыбы, зараженные паразитами, даже в начальных стадиях инвазии питаются более мелкими объектами и менее интенсивно.
· Интенсивность питания. Для каждого вида рыб характерны свои температурные границы, в которых питание происходит наиболее интенсивно, ослабевает или прекращается. Очень сильно влияет на интенсивность питания состояние рыбы — упитанная рыба питается менее интенсивно, чем истощенная: годовики карпа после зимнего голодания питаются гораздо активнее, чем сеголетки в конце лета. При определении интенсивности питания учитывают количество пищи, которое находится в пищеварительном тракте в данный момент, суточную ритмику питания и скорость продвижения пищи по тракту. Общее предварительное представление об интенсивности питания дает индекс наполнения пищеварительного тракта — отношение массы пищи, находящейся в пищеварительном тракте, к массе тела, выраженное или в процентах, или в продецимилле (отношение массы пищи к массе тела, выраженное в десятитысячных долях и обозначаемое 0 /000 ). Этот индекс характеризует накормленность рыбы в данный момент.
· Индекс наполнения может быть общим, если учитывается масса всего пищевого комка, или частным, если определяют долю какого-то компонента. Индексы потребления пищи, например, у личинок плотвы колеблются от 0 до 142%; на более поздних этапах максимальные индексы превышают минимальные в 3—10 раз.
· В течение суток рыбы питаются неравномерно. Промежутки активного питания чередуются с периодами ослабления или даже прекращения потребления пищи. Определенные суточные ритмы обнаруживаются уже на самых ранних этапах развития. На этапе смешанного питания периоды интенсивного поглощения пищи чередуются с периодами полного прекращения питания (ночью). В дальнейшем полного опорожнения кишечного тракта ночью не наблюдается, но интенсивность питания снижается.
· Суточная ритмика питания определяется биологическими особенностями самой рыбы и качеством и поведением пищевых объектов. У мирных рыб, особенно планктоноядных, перерывы в питании невелики, у хищных они могут длиться больше суток. Суточные ритмы питания многих карповых рыб имеют в большинстве случаев два максимума — утром и вечером. В неблагоприятных условиях, при резких сдвигах температуры, отсутствии пищи — суточные ритмы могут нарушаться. На интенсивности питания рыб отражаются также суточные миграции основных объектов питания — планктонных организмов, личинок хирономид, бокоплавов и т. д.
· Пищевой рацион. При изучении питания обычно пытаются установить пищевой рацион рыбы, т. е. количество пищи, съедаемой ею за какой-то период, выраженное в процентах от массы тела.
· Чаще всего определяют суточный рацион, но можно подсчитать и годовой. Существует прямой способ вычисления — определение количества съеденной пищи, но обычно пользуются косвенными способами, основанными на результатах азотистого баланса, потребления кислорода, учета индексов наполнения кишечника, скорости переваривания пищи и т.д. Например, суточный рацион можно определить по формуле D) = A(24/n), где D — суточный рацион, %; А — средний индекс наполнения кишечника, %; п — скорость переваривания пищи, ч.
· Пищевой рацион рыбы зависит от ряда биотических и абиотических факторов: ее физиологического состояния, интенсивности обмена, качества пищи, а также температуры, содержания кислорода.
· В частности, большое значение имеет качество корма. Сеголетки щуки в день съедают циклопов 160—175% массы тела, олигохет 150—330, личинок хирономид 150—250, рыбы — 30—50%Вобла съедает в сутки мизид 17% массы тела, а моллюсков—28%. Суточный рацион прудовых рыб составляет 1,5—8% массы тела. Значительное влияние на величину суточного рациона оказывает возраст рыбы: у молоди он выше, у взрослых — ниже (у личинок карпа в нерестовых прудах он составляет 20—60%, карпа-годовика — 6—8, у двухлетка — 2%).
· Физиологическое состояние рыбы отражается на интенсивности питания очень сильно. Истощенная рыба питается более активно, чем упитанная. У леща осенью при коэффициенте упитанности Кy = 2,3 индекс наполнения кишечника составлял 197 0/000, тогда как при Кy =2,5 он был вдвое меньше и не превышал 86 0/000 . Многие рыбы перестают питаться перед нерестом. Ослабляется питание во время заболевания и т.д.
· Имеет значение также видовая принадлежность рыбы. Молодь осетра ест больше, чем молодь севрюги. Суточные рационы тесно связаны с особенностями поведения рыб. В одиночку и в группе рыбы питаются неодинаково, при этом планктонофаги интенсивнее питаются в стае, бентософаги и хищники успешнее выслеживают добычу и овладевают ею поодиночке.
· Интенсивность питания рыб колеблется по сезонам: весной и осенью при одинаковой температуре она различна.
· Из физико-химических факторов среды первостепенное значение имеют температура и газовый режим. Каждый вид рыбы может питаться в определенных температурных границах, выше и ниже которых рыба не берет пищу (для карпа оптимальная температура 20—27°С; при температуре ниже 4°С и выше 35°С он не питается).
· При неблагоприятном газовом режиме величина суточного рациона снижается.
· При определении суточного рациона прудовых рыб необходимо учитывать также физиологическое действие кормов. Под действием различных кормов наступают изменения строения стенок кишечника, количества гемоглобина, интенсивности эритропоэза и лейкоцитарной формулы крови.
· Однообразные корма растительного происхождения, в частности льняной (бедный лизином) и соевый шрот (бедный метионином), вызывают наибольшие отклонения от нормы в строении кишечного тракта. Если принять за норму строение кишечника карпа, питавшегося естественной пищей, то у рыб, выращенных на льняном жмыхе и соевом шроте, высота складок слизистой в 10—5 раз меньше, цилиндрический эпителий этих складок или отсутствует, или полуразрушен и невысок, а продольный циркулярный слой гладких мышц сильно истончен.
· Разрушение цилиндрического эпителия должно повлечь за собой нарушение его физиологических функций — секретирования пищеварительных ферментов и всасывания питательных веществ.
· Показателем потребления и усвоения пищи является кормовой коэффициент (количество корма, необходимое для прироста единицы массы).
· Характер питания является видовым свойством рыбы, с питанием определенной пищей связаны особенности ее организации: органов чувств, ротового аппарата, пищеварительного тракта, обеспечивающих обнаружение соответствующих объектов, их схватывание и переваривание.
· Характер питания рыб одного вида, но разного возраста, пола, и разных видов, населяющих водоем, их взаимоотношения при использовании пищевой базы, т. е. их пищевые отношения, или пищевые связи, являются основой внутривидовых, межвидовых и вообще биотических связей в водоеме.
· Непосредственное представление о характере питания рыб дает анализ содержимого пищеварительного тракта (для безжелудочных рыб — кишечника): определив процентный состав пищевых организмов в пищевом комке, можно предварительно судить о значении для данной группы рыб различных пищевых объектов.
· Однако состав пищи рыб определяется не только наличием пищевых объектов, но и отношением к ним рыб.
· Рыбы относятся к пищевым организмам не одинаково, а более или менее избирательно, т. е. одних предпочитают, а других избегают.
· У планктоноядных рыб, которые захватывают сразу большое количество пищевых организмов, отфильтровывая их из значительного объёма воды, избирательная способность выражена слабо.
· У бентософагов, питающихся одиночными организмами, она проявляется довольно четко.
· Например, карп избегает остракод, хотя они крупнее других рачков и многочисленнее; севрюга выбирает мизид, стерлядь — хирономид.
· Способность рыб потреблять пищевые организмы в ином соотношении, чем они имеются в водоеме, получила название избирательной способности. Для количественного выражения ее был предложен соответствующий индекс, который представляет собой отношение процента данной группы пищевых организмов в пищевом комке к проценту этой группы в фауне водоема (А. А. Шорыгин, 1948):
ИС = r/B
где ИС индекс избирательной способности, r — процентное значение группы пищевых организмов в пище рыбы, В — процентное значение этой группы в окружающей среде
· При этом значение В определяют по пробам планктона (если определяют индекс избирания планктонных организмов) или бентоса (если определяют избирательность бентических организмов).
· Величина ИС может сильно колебаться — от нуля до нескольких сотен. Если рыба питается всеми организмами подряд, соотношение пищевых организмов в ее пищевом комке и водоеме будет одинаковым и индекс будет равен единице; если рыба предпочитает какой то объект и берет его чаще других, его в комке будет больше и индекс будет выше единицы; если рыба избегает каких-то организмов, их доля в пищевом комке уменьшится, следовательно, индекс будет меньше единицы. ИС меняется в зависимости от вида и состояния рыбы (возраст, пол) и богатства и доступности пищевой фауны, которая, в свою очередь, определяется сезоном, циклом развития пищевых организмов, климатическими условиями и характером водоема.
· Например, для бентосоядных рыб большое значение имеет возможность доставать пищевые организмы с разной глубины. Эта способность перерывать грунт неодинакова у разных видов, а у рыб одного вида сильно зависит от характера дна (рис. 40, 41).
Таблица 9 Состав пищевого комка двух видов карасей, % .
| Группа пищевых организмов | Золотой карась | Серебряный карась |
| Ветвистоусые рачки Веслоногие рачки Личинки хирономид Обрывки растений Прочие организмы | 24,2 9,6 45,0 14,0 7,2 | 39,0 16,2 9,0 - 6,5 |
· Рассматривая взаимоотношения разных видов, прежде всего важно выяснить, расходятся ли они в выборе пищевых объектов или их “запросы” совпадают.
· Для определения степени совпадения пищи двух групп рыб существует соответствующий показатель — индекс пищевого сходства. Его вычисляют, исходя из состава пищевого комка: выписав весовой состав пищи (в процентах) двумя параллельными столбцами, отмечают меньшие значения для организмов, встречающиеся у обоих видов рыб. Сумма этих меньших значений и составит индекс пищевого сходства.
· Если характер пищи двух групп рыб полностью совпадает, индекс пищевого сходства достигает 100%; если совершенно различен, индекс будет равен 0. Например, индекс пищевого сходства по зоопланктонным организмам золотого и серебряного карасей по данным табл. 9 составит 49,3% (меньшие значения выделены).
· Эти же показатели можно представить графически в виде двух пересекающихся кривых. Площадь, общая для обоих видов, соответствует сумме меньших значений организмов в пищевом комке, т. е. индексу пищевого сходства (рис. 42).
· Индексы пищевого сходства меняются в зависимости от состояния (возраста, пола и т. д.) рыб, сезона года, времени суток, состава пищевой базы.
· С уровнем пищевого сходства между группами рыб связана интенсивность их питания. При меньшем индексе, т. е. при большем расхождении в выборе пищевых объектов, интенсивность питания повышается. Если пищевое сходство велико, то быстро ощущается недостаток пищи, рост рыб замедляется и в конечном счете какая-то группа их будет подавлена. Так происходит в некоторых озерах, где сорная рыба, более многочисленная, вытесняет промысловую, поскольку основу питания и тех и других составляют общие пищевые объекты.
· В небольших замкнутых водоемах, где пищевые ресурсы ограничены, в более выгодном положении оказываются рыбы, обладающие широким спектром питания, так как они легко могут сменить пищевой объект, освоить свободную экологическую нишу и избежать тем самым обострения пищевых отношений с другими рыбами.
· Показателен пример с питанием плотвы. Пищевой спектр взрослых особей этого вида очень широк, поэтому, обитая в разных водоемах — от пресноводных до солоноватоводных, они используют в пищу преимущественно растения, планктон, или моллюсков в зависимости от того, какая группа меньше используется другими рыбами.
· Среди объектов тепловодного хозяйства широкий пищевой спектр свойствен серебряному карасю. Он успешно может потреблять низшие растения (фитопланктон), зоопланктон, бентос, детрит.
· При рыбохозяйственном освоении водоема товарным продуктом является рыба. Она — последнее звено преобразований органического вещества в водоеме. Цепь жизни в водоеме начинается с водорослей. Если расположить обитателей водоема в ряд по характеру их питания, то получим так называемую трофическую “пищевую цепь”, в которой каждое звено служит пищей следующего. В общем виде эта цепь отражает пищевые отношения трех больших групп гидробионтов: водные растения —> беспозвоночные (зоопланктон, зообентос, прибрежная фауна) —> рыба (первичная продукция) —> (промежуточная продукция) —> (конечная продукция)
· Организмы первой группы — растения — используют в процессе жизнедеятельности неорганические вещества (углекислоту, минеральные соли), т. е. образуют с участием солнечной энергии начальное звено органической жизни в водоёме.
· Детализация пищевых цепей зависит от конкретной задачи. Можно составить ее для каждого вида. У взрослых растительноядных рыб (белый толстолобик) эта цепь будет короткой: водоросли — рыба; у хищников — длинной: водоросли — зоопланктон — бентос — мелкая рыба — хищная рыба.
· При переходе с одного трофического уровня на другой теряется 80—90% энергии, а усваивается 10-20%. При удлинении пищевой цепи затраты энергии на получение конечной продукции (рыбы) многократно увеличиваются. Это связано с тем, что в каждом звене пищевой цепи прирост живой массы ниже затрат пищи в 5—10 раз и более. На образование 1 кг зоопланктона затрачиваются десятки килограммов фитопланктона; хищные рыбы на прирост 1 кг массы потребляют 5—10 кг мелкой рыбы и т. д. В результате в длинных пищевых цепях значительны весовые потери. В коротких пищевых цепях, т. е. при выращивании рыб детритоядных, растительноядных или планктоноядных, конечный продукт — рыба — образуется быстрее и в большем количестве и продуктивность водоема увеличивается.
· Таким образом, мирные рыбы, способные питаться детритом и растительностью, выгодны в том отношении, что они используют эту пищу непосредственно на построение собственных тканей, т. е. конечного продукта, а не в качестве промежуточных продуктов, как например, бентосоядные рыбы — бентос.
· Заселение водоема рыбами, различающимися по характеру питания, способствует более полному использованию различных звеньев общей пищевой цепи и повышению продуктивности водоема в целом. Поэтому в прудовом рыбоводстве большое значение имеет подбор видов для поликультуры применительно к характеру водоема (донные отложения, флора, беспозвоночные, рыбное население и т. д.).
· В естественных водоемах изменения характера пищи в течение жизни рыбы обеспечивают расхождение в питании видов, имеющих даже близкие пищевые спектры (многие карповые), а внутри одного вида — у рыб разного возраста, размера, пола и т. д., благодаря чему ослабляется напряженность пищевых отношений. Однако это равновесие может быть нарушено в результате хозяйственной деятельности человека. Нарушение пищевых отношений вызывает изменение состава стада рыб (популяции). В больших водоемах результат сказывается медленнее, в малых — значительно скорее. При перелове одних видов их численность сокращается, но одновременно нередко усиленно развиваются другие виды. Перенаселение водоема ведет к уменьшению его продуктивности, так как сопровождается измельчанием рыбы и замедлением ее роста.
· Учитывая, что основные закономерности пищевых отношений — и межвидовые и внутривидовые — в значительной степени определяют динамику численности и биомассы стада (популяции) рыб, в интересах рационального рыбного хозяйства важно воздействовать на них в нужном направлении.
· Жирность и упитанность рыб. С условиями и характером питания в течение года, а также с ростом, возрастом и полом рыб тесно связано содержание жира в теле (жирность), а значит питательная ценность рыб как пищевого продукта. При этом сезонные колебания жирности яснее выражены у половозрелых рыб по сравнению с неполовозрелыми, а у самок заметнее, чем у самцов.
· У одних видов отложение жира четко локализовано: в мышцах (лососи), в печени (акула, треска), в полости тела между органами (судак). У других видов локализация жира менее выражена (осетровые, сельди).
· По содержанию жира в мышцах рыб разделяют на четыре группы: тощих (0,2—1,2% жира — щука, бычки, навага, треска, окунь, судак), среднежирных (1,5—4,5%—вобла, камбала, лещ, сазан), жирных (5—15%—лососи, осетровые, скумбрия) и особенно жирных (более 15% — угорь, минога, хамса).
· Если на протяжении года условия питания вида колеблются сильно или годовые ритмы включают периоды голодания (зимовки, миграции), то в теле рыб сезонно накапливаются большие запасы жира, которые расходуются в периоды перерыва в питании. Если условия питания в течение года меняются мало, а больших передвижений рыбы не совершают, то их жирность в течение года изменяется мало.
· Для определения упитанности рыб кроме данных химического анализа удобно пользоваться формулой Фультона в модификации ВНИИПРХа, основанной на положении, что упитанность связана с объёмом тела, его плотностью и жирностью:
Ку=Р*100/L3
где Ку — коэффициент упитанности; Р — масса тела, г; L — длина тела, см.
· Однако в формуле Фультона из общей массы тела не исключается масса гонад и содержимого пищеварительного тракта, которые могут быть значительными, но не имеют отношения к упитанности. Ф. Кларк считает, что для более точного определения упитанности следует учитывать массу тела без внутренностей.
· У большинства карповых, лососевых и осетровых рыб показатели жиронакопления очень неустойчивы (табл. 10). У таких рыб, как щука, судак, они не претерпевают значительных колебаний.
· В разных частях и органах тела рыбы жир накапливается и расходуется неравномерно в зависимости от видовой специфики, условий питания.
Таблица 10 Динамика коэффициента упитанности карпа /
| Возрастная группа | Масса, г | Ку |
| Личинки | 0,010-0,026 | 0,8-1,1 |
| Мальки | 0,7-5,1 | 1,1-3,5 |
| Сеголетки | 25,0-28,0 | 1,9-3,39 |
| Годовики | 20,0-22,0 | 1,7-2,31 |
| Двухлетки | 500,0-600,0 | 2,6-3,2 |
| Производители | 2800,0-8000,0 | 2,4-3,4 |
· При недостаточном питании линейный рост прекращается, но накопление жира продолжается, поэтому . даже в неблагоприятных условиях у взрослых рыб обеспечивается развитие гонад. У голодающих рыб жир гонад расходуется в последнюю очередь, поэтому даже при истощении родителей потомство бывает обеспечено питательными веществами (жир в икринке).
· У наиболее выносливых рыб нашей фауны — карасей — выработалась способность перестраивать обмен в период зимовки таким образом, что к концу зимнего голодания количество жира у них в теле не уменьшается. Предполагают, что это необычное явление связано с процессами анаэробного гликолиза и превращения части углеводов непосредственно в жир.
· Значение запасов жира особенно возрастает у рыб, не питающихся зимой, в частности прудовых карповых (карп, карась). В период зимнего голодания жир, с одной стороны, служит источником энергии в процессах обмена веществ, а с другой — образует тепловую защиту внутренних органов и тканей и таким образом регулирует тепловой обмен в условиях низких температур. При этом степень изоляции внутренних органов жировой тканью различна у рыб разной массы: у крупных она больше, чем у мелких. В результате, например, у нестандартных сеголетков расход зимой резервного жира ведет к ослаблению тепловой изоляции головы и спинного мозга, что является одной из причин потери ориентации, нарушения локомоторной функции и в конечном счете гибели. Запасы жира служат также источником энергии для развития гонад. Однако чрезмерное накопление жира вредно, так как вызывает нарушение обмена веществ.
Рост и возрастная изменчивость рыб
· Познание закономерностей роста рыб, а в естественных водоемах возрастного состава стада дает представление о степени использования рыбой пищевой базы, позволяет определить время и размеры наиболее целесообразного вылова и т. д. Иначе говоря, оно дает основание для составления прогнозов о составе стада и масштабе промысла, а в прудовом рыбоводстве является основой для племенной работы, для установления наиболее выгодных с хозяйственной точки зрения сроков выращивания рыбы.
· Продолжительность жизни рыб и их размеры весьма различны, но специфичны для каждого вида. Самые маленькие рыбки — бычки с Филиппинских о-вов — не превышают в длину 7,5 — 14,0мм, продолжительность их жизни — около года. В тропиках в пресных водах живут также крошечные представители семейства Cyprinodontidae (карпозубых). Другая крайность — белуга массой около 1,5т, возраст которой может составить 100 лет, или полярная акула, достигающая в длину 15 м и даже больше. Обыкновенный сом достигает массы 300 кг. 100 лет, конечно, редкость. Обычно предельный возраст “долгожителей” исчерпывается 25—30 годами. У наших пресноводных рыб предельный возраст меньше: у аральского леща — 15 лет, амурского серебряного карася — 12, амурского сазана — 16, карпа — 20 лет.
· Величина рыб одного вида и одной возрастной группы из разных водоемов может различаться в зависимости от условий жизни. Например, в Подмосковье зарегистрированы сеголетки карпа массой 200 г, а на о-ве Ява — трехмесячные карпы массой 690 г.
· В настоящее время в связи с интенсификацией промысла лишь ограниченное количество особей доживает до предельного видового возраста. Естественная продолжительность жизни определяется видовыми особенностями обмена веществ. Многие виды рыб погибают после первого нереста (дальневосточные лососи рода Oncorhynchus, угорь Anguilla). Некоторым видам свойственна частичная посленерестовая гибель (у благородного лосося — семги — после нереста гибнет до 87% особей, причем преимущественно самцов).
· Рост неразрывно связан с развитием, являясь одной из его сторон. Специфической особенностью роста рыб является преобладание ассимиляции над диссимиляцией, благодаря чему рыба растет в течение всей жизни и после наступления половой зрелости. Характерным свойством рыб является снижение обмена (но без нарушения функций органов) при длительном недостатке или отсутствии пищи и быстрое восстановление его интенсивности при улучшении условий питания. Эта способность позволяет многим рыбам безболезненно переносить долгое зимнее голодание.
· Различают весовой рост (наращивание массы тела) и линейный (увеличение длины тела). Весовой рост сильнее подвержен колебаниям в зависимости от условий питания, чем линейный, поэтому закономерности изменения роста, его специфику легче проследить на линейном росте. Вместе с тем в прудовом рыбоводстве основным показателем эффективности выращивания рыбы является весовой рост. Рыба растет неравномерно как в течение жизни, так и в течение всего года, причем в разные периоды рост ее характеризуется определенными особенностями.
· Прежде всего различен характер роста рыб до и после наступления половой зрелости. Обычно до наступления половой зрелости рыбы растут наиболее быстро. Пища используется ими в основном на весовой и главным образом линейный прирост, т. е. является в основном продуцирующей. Поэтому в первые годы жизни, как правило, происходит наиболее быстрое нарастание линейных размеров. После наступления половой зрелости темп роста снижается, но наращивание массы тела продолжается и максимальное увеличение ее наблюдается именно в старшем возрасте. Значительная часть потребленной пищи расходуется на образование половых клеток и накопление резервных веществ, обеспечивающих созревание гонад, благополучный исход зимовки, миграции и т. д. Доля продуцирующей пищи уменьшается и соответственно увеличивается часть пищи, идущей на поддержание жизнедеятельности (поддерживающая пища).
· В период старения организма линейный рост сильно замедляется. Пища расходуется в основном на поддержание жизненных процессов. Такова общая закономерность, отражающая характер развития рыб в течение жизни. В ее пределах рост рыб в отдельные, более короткие, промежутки времени определяется многими факторами: температурой, гидрохимическим режимом, обеспеченностью пищей, состоянием здоровья, особенностями обмена на разных этапах развития, полом, видовой принадлежностью и т. д.
· Неравномерность роста зародышей карпа в икринке в период инкубации, т. е. до выклева (в нерестовом пруду хозяйства ТСХА), представлена на рис. 43. Из рисунка видно, что более быстрое нарастание массы охватывает дробление, обрастание желтка, начало формирования органов зародыша, т. е. время усиленного формирования и дифференцировки тканей. После закладки органов следует сильное замедление весового роста. Резкий подъем последнего отрезка кривой показывает быстрое увеличение массы зародыша перед выклевом.
· Рост молоди в начальный период жизни после выклева также происходит неравномерно. Именно в первые 25—30 дней после выклева выявляются четыре цикла, в которых повышенная скорость роста чередуется с замедленной (табл. 11).
Таблица 11 Периодичность роста молоди карпа в нерестовых прудах (ТСХА)
| Возраст молоди , дни | Длина тела, мм | Масса Р, мг | Кратность увеличения массы | Цикл развития |
| 1 | 5,5 | 1,2 | ||
| 2-3 | 5,5 | 1,2 | ||
| (переход на питание) | 6,8 | 1,5 | 5 | I |
| 6 | 8,0 | 6,1 | II | |
| 8 | 10,0 | 16,3 | 2,7 | II |
| 12 | 13,5 | 30,0 | III | |
| 18 | 20,0 | 150,0 | 5 | III |
· I цикл—от вылупления личинок до 4—6-дневного возраста; характеризуется наиболее высокой интенсивностью роста и питания. Биологической особенностью этого цикла является зависимость основных функций организма — питания, дыхания — от желточного мешка, поставляющего организму пищу и кислород.
· II цикл—от 4—6-дневного до 6—10-дневного возраста; характеризуется значительной депрессией дыхания и роста. Резорбция желточного мешка вместе с его кровеносными сосудами приводит к понижению функций дыхания и эндогенного питания. Газообмен происходит с помощью личиночных органов. К активному питанию личинки еще не приспособились, они питаются неинтенсивно и в той или иной степени испытывают недостаток пищи. За это время масса тела увеличивается примерно в 2,7 раза.
· III цикл — от 10-дневного до 18—20-дневного возраста — новый подъем дыхания и роста. Сформированы жабры, органы пищеварения, передвижения (плавники) и т. д. Увеличение массы за это время — примерно в пять раз.
· IV цикл — от 18—20-дневного до 27—30-дневного возраста; заканчивается формирование личинки в малька, появляется чешуя. При обильном питании скорость роста сохраняется высокая, при скудном корме — резко снижается.
· Из этих данных следует, что циклы III и IV, характеризующиеся повышенной скоростью роста, являются очень ответственными в раннем развитии рыб. Обильное кормление на этих стадиях обеспечивает хороший рост, а скудное питание приводит к общей депрессии роста. Поэтому нужно пересаживать молодь из нерестовых прудов в выростные во время цикла II, пока рост замедлен, т. е. до 8-дневного возраста, чтобы цикл III проходил уже в условиях обилия пищи. В этом случае будет использована высокая энергия роста организма, способность его к быстрому росту в это время.
· У рыб, как у животных с непостоянной температурой тела, наблюдается периодичность, неравномерность роста в течение года. Периоды усиления и замедления роста чередуются в течение года и сезона в соответствии с наиболее или наименее благоприятными для данного вида температурными условиями, богатством или бедностью пищевой базы и т. д. У неполовозрелых карповых, сиговых и многих других рыб характер роста наиболее ясно отражает обеспеченность их пищей и, следовательно, подвержен резким колебаниям. У хищных, например окуневых, рыб зависимость роста от обеспеченности пищевыми организмами выражена слабее. Это связано с тем, что в случаях обеднения пищевой базы они поедают себе подобных (каннибализм).
· Очень специфичен рост рыб, которые перестают питаться в период зимовки (большинство объектов разведения в тепловодных хозяйствах) или миграций (проходные рыбы): у них перед зимним голоданием или длительным странствованием в тканях тела накапливается жир при замедленном белковом росте. При этом обычно после зимовки, в начале вегетационного периода, увеличения линейных размеров тела не происходит, а преобладает накопление жира. Этот процесс продолжается до момента достижения определенной упитанности. После этого жиронакопление замедляется или приостанавливается, а ускоряются белковый рост и связанный с ним линейный прирост.
· Таким образом, очень важным показателем качества посадочного материала, например карпа, является его упитанность, в какой-то мере отражающая уровень накопления резервных веществ, а следовательно, и подготовленности рыбы к зимнему голоданию и дальнейшему росту во второе лето. Чем упитаннее годовики после зимовки, тем скорее начинается увеличение их линейных размеров после посадки в нагульные пруды.
· Рост рыбы в значительной мере отражает качество и количество Пищи, находимой ею в водоеме. Поэтому, как правило, в течение Вегетационного периода увеличиваются различия в величине молоди, полученной одновременно от одних родителей. Например, при выращивании сеголетков карпа разница в размерах выклевывающейся молоди не превышает нескольких процентов, а осенью, т. е. в конце первого лета жизни, самые крупные экземпляры превосходят по массе самые мелкие зачастую в 10—20 раз. Возрастает изменчивость по массе рыб к осени и в нагульных прудах. При этом чем большие колебания размеров наблюдаются при зарыблении прудов весной, тем более разнокачественной оказывается рыба в конце выращивания.
· Условия питания определяют быстроту индивидуального роста рыб, а тем самым и характер роста стада в целом. Богатая кормовая база обеспечивает быстрый рост. При этом все рыбы могут потреблять одинаковую пищу, поэтому они растут относительно ровно и индивидуальная изменчивость в стаде невелика.
· При недостатке кормовых организмов скорость роста стада замедляется. Даже небольшие различия в исходной величине позволяют рыбам питаться по-разному и различия в росте увеличиваются, становятся все более заметными, а индивидуальная изменчивость рыб в стаде повышается. Таким образом, рост является обобщающим показателем того, насколько условия обитания отвечают потребностям рыбы.
· Однако реакция организма на изменение условий жизни неоднозначна. В условиях обильного питания наряду с ускорением роста повышаются, например, выживаемость, особенно молоди, плодовитость, увеличивается численность вида; обеднение пищевой базы, замедление роста рыб сопровождаются уменьшением плодовитости, нередко проявлением каннибализма и в конечном счете уменьшением численности вида.
· Неравномерный в течение года рост рыб (сезонные ритмы) приводит к образованию слоев (колец) на чешуе и костях. Эту особенность используют для определения возраста и расчисления темпа роста рыб в предыдущие годы, для чего исследуют чешую, отолиты, отдельные кости скелета — жаберную крышку, лучи плавников, клейтрум, позвонки и т.д. (рис. 44).
· У большинства рыб, в том числе карпа, возраст и рост определяют по чешуе. Для приготовления препарата берут несколько чешуек со средней части тела между основанием первого спинного плавника и боковой линией, промывают в слабом растворе нашатырного спирта (1 —10%), обсушивают мягкой тканью и помещают между двумя предметными стеклами. Наилучшие препараты получаются из чешуи, взятой от свежей рыбы, но можно использовать соленую и фиксированную в формалине.
· У осетровых, сомов, судака и некоторых других, как правило, используют луч грудного плавника, поперечный срез которого подшлифовывают и приклеивают бальзамом на предметное стекло.
· У окуневых, налима и некоторых других рыб удобны для определения плоские кости — жаберные крышки и клейтрум. Косточку очищают, обваривая кипятком, обезжиривают в смеси эфира и бензина в отношении 1:2 и подсушивают. Формалинный материал не используют. Так же подготавливают позвонки, которые раскалывают продольно и рассматривают слом.
· У тресковых, камбаловых и судака хорошо определять возраст по отолитам. Отолиты вынимают из свежей рыбы, чаще всего разрезав голову вдоль по середине или поперек в области затылка, разламывают пополам, подшлифовывают и прокаливают. На шлифе хорошо видны годовые кольца.
· Для большей достоверности результатов рекомендуют определять возраст рыб параллельно по чешуе и костям. На чешуе и костях видны расположенные друг за другом светлые и темные полосы, или кольца, которые отражают рост рыбы в течение года. Полосы, образующиеся в периоды замедленного роста рыбы, состоят из мелких клеток, плотно прилегающих одна к другой; в падающем свете (освещение сверху) полосы темные, в проходящем (освещение снизу) — светлые. В периоды усиленного роста клетки, образующие полосу, крупнее и лежат свободнее. Кольцо оказывается более широким, матово-светлым в падающем свете и темным — в проходящем.
· В умеренных широтах наиболее интенсивное питание и, следовательно, быстрый рост рыб приходится на лето или даже осень, в арктических районах — на зиму, что соответственно отражается на ширине колец. У рыб, обитающих в тропических и экваториальных водах, на костях и чешуе также откладываются сезонные кольца, иногда они соответствуют периодам засухи и тропических ливней.
· Широкая и узкая полосы составляют вместе одну годовую зону (рис. 45).
· Таким образом, узкие и широкие полосы на чешуе и костях появляются вследствие изменения скорости роста тела. Неправильно называть узкие темные кольца “зимними”: если зимой рыба не питается (карп и др.), то она и не растет и закладки кольца не происходит. Эти кольца соответствуют периодам медленного роста, они могут закладываться весной, осенью и даже летом.
· В связи с этим на чешуе и костях могут образовываться не только годовые кольца, но и так называемые дополнительные (добавочные), появляющиеся в результате ослабления или прекращения питания в преднерестовый или нерестовый период, перемены кормовых объектов и т. п. К добавочным кольцам относится прежде всего первое кольцо на чешуе — мальковое, которое возникает при переходе молоди с питания планктоном на питание бентосом. Это кольцо оказывается внутри первой годовой зоны.
· У многих рыб, не питающихся во время нереста, этот перерыв в питании отражается на чешуе в виде так называемых “нерестовых” колец (марок), которые обнаруживаются по разрушенному, как бы размытому краю.
· Из-за различия в характере роста рыб до и после наступления половой зрелости у разных возрастных групп время закладки годовых колец сдвигается. Например, у неполовозрелых особей азовского судака и аральского леща, растущих наиболее быстро, годовые кольца закладываются в начале вегетационного периода — еще весной. После наступления половой зрелости определенная доля пищи расходуется на созревание гонад. Поэтому у рыб старших возрастных групп в начале вегетационного периода, когда интенсивно накапливаются резервные вещества, линейный рост часто не происходит. Через определенный промежуток времени жиронакопление замедляется или приостанавливается. Тогда ускоряется линейный рост и начинает закладываться новое широкое кольцо, это происходит уже во второй половине лета.
· Таким образом, для правильного “чтения” чешуи при определении возраста необходимо знать биологию рыб и прежде всего особенности их роста.
· Для обозначения разных возрастных групп принята следующая терминология (знак “+” обозначает прирост следующего года):
| Возрастная группа | Число колец | Обозначение |
| Сеголетки | нет | 0+ |
| Годовики | Одно | 1 |
| Двухлетки | Одно | 1+ |
| Двухгодовики | Два | 2 |
| Трёхлетки | Два | 2+ |
| Трёхгодовики | Три | 3 |
· Пригодность темных и светлых полос на чешуе и костях рыб для определения возраста открыл еще Левенгук (1684), отметив, что их возникновение связано с неодинаковой быстротой роста. Если знать соотношение роста чешуи (или костей) и роста тела, то можно восстановить линейный рост рыбы в годы, предшествующие поимке (обратное расчисление роста).
· Норвежский исследователь Э. Леа на основании допущения, что длина тела рыбы и длина чешуи изменяются в прямой пропорции, т. е. находятся в прямолинейной зависимости ln : l = Vn :V, предложил формулу ln=Vnl /V, где l — длина тела рыбы в момент поимки, V— длина чешуи по средней линии от центра до края, 1n — вычисляемая длина рыбы в возрасте п лет, Vn — расстояние от центра чешуи до годового кольца в возрасте п полных лет.
· Установленная зависимость иллюстрируется на рис. 46. Чешую проецируют рисовальным аппаратом на лист бумаги или (в модификации Г. Н. Монастырского) на специальную доску.
· Зная длину l рыбы в предыдущие годы, можно вычислить ее еже годные приросты (t): l1, = l1,, t2 = l2—l1 и т. д. Данный метод дает сравнимые результаты.
· Впоследствии в положения, выдвинутые Э. Леа, были внесены по правки и уточнения; было показано, что между ростом тела и чешуи у некоторых рыб существует не прямая, а логарифмическая зависимость: пропорциональны лишь приращения логарифмов длины тела и длины чешуи. Специальный прибор (Г. Н. Монастырского) позволяет расчислить рост методом логарифмических шкал. Характер образующихся кривых сходен с данными, полученными методом Леа.
· При анализе роста рыб пользуются различными показателями в зависимости от поставленной задачи. Обычно вычисляют:
· а) весовой или линейный прирост: W1,—W0 (W1—конечная величина, W0 — начальная величина);
· б) относительный прирост, или темп роста: (W1,—W0 )/W0 (обо значения те же);
· в) относительную скорость роста К (показывает рост в определённый промежуток времени):
к = Wt - Wt /[(Wt + W0 )/2] * t
где W0 величина тела в начале периода, Wt — величина тела в конце периода, t — промежуток времени.
· Рост рыб взаимосвязан с возрастом: рыбы могут расти практически всю жизнь. Однако, как и у всех живых организмов, с возрастом у них происходят изменения в молекулярной структуре клеток, в соотношении белков, нуклеиновых кислот, липидов, в количестве и качестве ферментов, в способности образовывать антитела. Это существенно влияет на энергетический и пластический обмен всего организма, поэтому его жизнедеятельность и прежде всего очень важные с хозяйственной точки зрения его приспособительные возможности и продуктивность в разном возрасте неодинаковы.
· Общие закономерности возрастной изменчивости рыб очень сильно отражаются и на их воспроизводительной способности: во второй половине жизни постепенно снижается плодовитость, а затем, в результате дегенерации гонад, прекращается способность к размножению. Этот процесс сопровождается изменениями качества половых продуктов и свойств получаемого потомства. Поэтому, как и в других отраслях животноводства, в прудовом рыбоводстве большое значение придается возрастному подбору производителей, который в значительной степени определяет формирование качества посадочного материала и затем продуктивность товарной рыбы.
· При изучении возрастных изменений основного объекта тепловодного хозяйства (ТСХА) — карпа — было обнаружено, что у производителей разного возраста качество половых продуктов различно. Эти различия касаются количества сухого вещества, белков, жиров, микроэлементов, показателей энергетического обмена. С самого раннего, эмбрионального, периода развития проявляются различия в качестве потомства.
· У самцов, созревающих впервые, сперма отличается наименьшим количеством сухого вещества, жира и золы, низким содержанием фосфорных соединений и нуклеиновых кислот, пониженной концентрацией и наименьшим количеством жизнеспособных сперматозоидов в семенной жидкости.
· У самцов среднего возраста в сперме отмечено высокое содержание сухого вещества, белков, жиров, аминокислот: цистина, цистеина, триптофана, гистидина; возрастание количества меди и уменьшение цинка. Сперма характеризуется большими запасами нуклеиновых кислот и энергетических веществ (лецитин, холестерин), что свидетельствует о высокой интенсивности обмена. Количество жизнеспособных спермиев в семенной жидкости и общая концентрация спермы наибольшие по сравнению с рыбами других возрастов.
· Сперма старых рыб имеет наибольшее количество жира, лецитина, наименьшее — меди, цинка, РНК, пониженное содержание АТФ; понижена также концентрация спермы.
· По мере увеличения возраста самок изменяется величина икринок: сначала она возрастает, а затем уменьшается или изменяется незначительно. Меняется и соотношение икринок разных размерных групп: количество крупных икринок сначала увеличивается, а потом уменьшается (табл. 12).
Таблица 12 Изменение величины икринок по мере увеличения возраста двух самок карпа
| Показатели |
Возраст самок, лет
|
||||||||||||||
|
|
| Количество, %, икринок разного диаметра, мм: | ||||||||
|
|
Икра средневозрастных самок содержит наибольшее количество сухого вещества, жиров, протеина и таких аминокислот, как цистин, цистеин, триптофан, гистидин и т. д.; в ней наибольшее количество марганца, меди и цинка.
· Характер нереста и основные показатели качества потомства при нересте производителей разного возраста неодинаковы: впервые нерестующие производители нерестуют обычно вяло и недолго (около 3 ч) или, наоборот, очень долго, с промежутками, в общей сложности больше суток. Икра отличается наименьшим оплодотворением (61 — 88,8%), наибольшим отходом в период инкубации, доходящим иногда до 100%, большей продолжительностью инкубации. Выход молоди от одной самки из нерестовых прудов крайне низок и неравномерен (0—90 тыс. шт.). Замедленный рост и большие отходы характеризует их потомство и при дальнейшем выращивании, в первое и второе лето жизни.
· У производителей, нерестующих второй раз, нерест длится дольше, он более активный. Оплодотворённость икры повышается (93,3%). Отход икры ниже, чем при первом нересте, но все же высок (14,3%). Выход молоди от одной самки из нерестовых прудов 31 —135 тыс. шт.
· Производители среднего возраста нерестуют бурно, в течение 5—8 ч. Икра оказывается наиболее оплодотворенной (91—98%), эмбриональное развитие потомства проходит быстро.
· У производителей среднего возраста отмечен наименьший отход икры, в ней больше жиров и белка, выклев молоди и переход ее на внешнее питание проходит быстрее. От каждой самки из нерестовых прудов получают наибольшее количество молоди (свыше 150 тыс. шт.). В постэмбриональный период эта молодь сохраняет преимущества по основным рыбоводным и биохимическим показателям (отходы, рост, накопление жиров, прирост белка и т. д.).
· Так относительное увеличение массы зародышей при инкубации икры в зависимости от возраста производителей выглядит следующим образом:
|
Возраст производителей, лет1 |
Относительное увеличение массы (Р2—Р1)/Р21 |
Возраст производителей, лет1 |
Относительное увеличение массы (Р2—Р1)/Р21 |
| 6 | 0,53 | 11 | 0,52 |
| 7 | 0,28 | 8 | 0,40 |
| 10 | 0,83 | 12 | 0,30 |
1 Икру производителей возраста 6 и 10, 7 и 11, 8 и 12 лет инкубировали одновременно (попарно). 2 P1, и Р2 — начальная и конечная массы зародыша.
· Стареющие производители нерестуют менее бурно, долго, с промежутками. Эмбриональное развитие молоди длится дольше, отходы икры повышаются (19—34%), весовой рост ее во время инкубации и желточного питания идет медленнее. Выклев и переход на активное питание растягивается. Выход молоди от одной самки из нерестовых прудов невысок (67—81 тыс. шт). При дальнейшем выращивании молодь характеризуется повышенным отходом.
· При дальнейшем старении карпов способность их к размножению затухает: некоторые пары перестают нерестовать в возрасте 13—15 лет. Повышаются отходы икры, молодь отстает в росте.
· По основным рыбоводным показателям — выживаемости, росту, зимостойкости — лучшие сеголетки и годовики получаются от производителей среднего возраста, худшие — от впервые нерестующих, поэтому рыбопродуктивность при выращивании потомства от производителей разного возраста различна.
Миграции и методы их изучения
· Число видов рыб, живущих вполне оседло, в одном месте, невелико (представители коралловых рыб, некоторые бычки семейства Gobiidae и др.). У большинства рыб обязательным звеном в жизненной цепи являются миграции — периодические сезонные перемещения в определенном направлении и в определенные сроки.
· Миграции могут происходить пассивно когда мигранты без собственных энергетических затрат подхватываются водными течениями в районе нереста и переносятся в другие районы. Так перемещаются пелагические икра и личинки, а иногда и неокрепшая молодь тех рыб, у которых места нереста и нагула разделены большим расстоянием. Например, у атлантической трески, нерестующей около Лофотенских о-вов (Скандинавия), пелагические икра и личинки переносятся в Баренцево море; у угря личинки с нерестилищ в Саргассовом море (Атлантика в районе Бермудских и Багамских о-вов) переносятся к берегам Европы.
· Большинству рыб свойственны активные миграции, когда передвижение совершается по определенному пути и, следовательно, сопровождается тратой энергии, зачастую очень значительной. Это передвижения к местам нагула или нереста трески, кефали, скумбрии, многих каспийских сельдей, дальневосточных лососей, угря и т. д. В зависимости от направления движения различают миграции анадромные (в реке вверх, против течения; в море — из глубин или открытой части в прибрежные районы) и катадромные (в реке — вниз по течению; в море — из прибрежных районов в открытое пространство). Например, миграции из моря дальневосточных лососей для нереста в верховья рек или каспийских сельдей и осетровых в Волгу, так же как трески из открытого моря к побережью, являются анадромными, а выход для нереста речного угря из рек в океан или отход морской камбалы в глубокие соленые участки моря — катадромными.
· Основными типами миграций являются нерестовые (от мест зимовки или откорма к нерестилищам), нагульные, или кормовые (от нерестилищ или зимовки к местам откорма), и зимовальные (из районов откорма или нереста в районы зимовки).
· Как видно из названий, каждый из этих типов миграций обусловлен определенными требованиями организма в тот или иной период жизненного цикла. Поэтому начало миграций определяется как внешними факторами (температура, течения), так и подготовленностью организма к изменению условий жизни — упитанностью, содержанием жира, состоянием зрелости гонад.
· Наиболее резкие изменения на протяжении миграций претерпевают проходные рыбы. Они резко меняют среду (морскую на пресноводную или наоборот), преодолевают большие расстояния и препятствия (семга проходит 1100—2500 км при скорости 50—100 км/сут., совершает прыжки при подъеме через пороги). У некоторых видов — дальневосточных лососей, угря, сельди-черноспинки — изменения в организме к концу нерестового хода становятся настолько значительными, что затрагивают структуру ДНК и делаются необратимыми. Вследствие этого рыбы в массе погибают после первого и единственного в жизни нереста.
· Пути полупроходных рыб значительно короче — из предустьевых пространств моря на нерестилища, расположенные в нижнем или среднем течении реки. Такие полупроходные рыбы, как судак, сазан, лещ, сом и др., в Волге с наступлением холодов перестают питаться и массами концентрируются, “залегают” на ямах дельты, где в оцепенении проводят зиму; с наступлением весны они покидают ямы и направляются к нерестилищам, расположенным в низовьях реки.
· Кроме морских, проходных и полупроходных рыб миграции свойственны многим пресноводным рыбам, но протяженность их передвижений невелика и не сопровождается столь значительными изменениями организма, которые вызвали бы посленерестовую гибель.
· Среди пресноводных рыб, помимо обитающих постоянно в одном месте, некоторые обитатели озер поднимаются для нереста в реки. Волховский сиг Coregonus lavaretus baeri до постройки гидростанции проходил из Ладожского озера через Волхов и озеро Ильмень в реку Мсту, а байкальский омуль С. migratorius — из озера Байкал в В. Ангару и Селенгу. Для нереста поднимается выше по реке, например, стерлядь. Карповые рыбы весной и в начале лета чаще передвигаются вверх по рекам, осенью — вниз по течению.
· Выяснение закономерностей миграций промысловых рыб очень важно для организации промысла. Наиболее надежным способом изучения миграций служит мечение рыб, позволяющее вполне достоверно установить пути, скорости движения рыб, скорость роста и т. д.
· Индивидуальные метки прикрепляют к жаберным крышкам, основанию спинного или хвостового плавников. Это делают специальными щипцами, леской, ниткой или проволокой. Метки изготавливают из нержавеющего материала, они могут быть в форме пластинки, диска, гидростатического цилиндрика и т. д., на которых нанесены номера и другие исходные данные. Метки должны быть хорошо заметными, минимально беспокоить рыбу и не вредить ей. Ультразвуковые метки позволяют следить за рыбой непрерывно в течение определенного времени.
· В целях изучения поведения больших групп рыб используют средства массового однородного мечения. К ним относят подрезание определенных плавников, окрашивание (как подкожное, поверхностных участков тела, так и тотальное), радиоактивное воздействие, применение антибиотиков — тетрациклина и др., после обработки которыми (инъекции или выдерживание в растворе) кости и чешуя при ультрафиолетовом облучении светятся желтым светом, и т. д.
· В рыбоводстве часто возникает необходимость мечения рыб, как индивидуального — при бонитировке производителей, так и группового.
· Распространено мечение путем подрезания плавников (предпочтительнее подрезать парные и прежде всего брюшные, так как другие быстрее восстанавливаются); с помощью цветных ниток, привязываемых к плавникам (на короткое время); таврения — выжигания метки на теле рыбы раскаленным или переохлажденным в жидком азоте клеймом и т. д. Метки, прикрепляемые к плавниковым лучам или жаберным крышкам; быстро теряются рыбами и при этом сильно травмируют их, особенно в летних прудах, когда рыбы неизбежно попадают в заросли. Применяют для мечения рыб и красители. Использовать красители можно разными способами. При прибавлении красителей к корму оказываются помеченными не только рыбы, получившие краску, но и их икринки и личинки. При купании рыб в красящих растворах окрашивается целиком поверхность тела, однако краска относительно быстро теряется. При индивидуальном мечении краску вводят под кожу с помощью инъекций. В этом случае метки сохраняются долго (несколько лет). В целях унификации чтения меток рекомендуют определенную систему выбора красок и места их введения. Например, для группового мечения рыб, разных по происхождению, краситель определенного цвета вводится одной группе под кожу жаберной крышки, а другой — под спинной плавник. Для индивидуального мечения производителей принята следующая система обозначения цифр и введения растворов разного цвета в разные участки брюшка: единицы обозначают синим цветом, десятки — красным, сотни — оранжевым; по средней — продольной линии брюшка располагают нечетные цифры, у основания плавников четные.
Место рыб в водных биоценозах
· Рыбы в водоеме вступают со всеми другими обитателями его в самые различные отношения. Эти отношения возникают как между рыбами одного вида (внутривидовые связи) и различных видов (межвидовые связи), так и между рыбами и представителями всех других систематических групп. Многообразные связи образуются при питании (связи потребителя и пищи, хищника и жертвы, паразита и хозяина и др.), при защите от врагов (образование стай, защита гнезд и молоди, покровительственная окраска), при размножении (связь представителей разных полов, родителей и потомства, охрана территории, выбор субстрата в период икрометания).
· Биотические и абиотические связи обитателей водоема переплетаются между собой. В результате этого сложнейшего процесса вырабатывается единство организма со средой обитания.
· Внутривидовые отношения рыб, формирующиеся в процессе эволюции, обеспечивают существование вида. Важным поведенческим элементом рыб одного вида является образование различных группировок. Основные из них — стая, скопление и стадо. Стая (косяк) — относительно продолжительная группировка, характеризующаяся сходством поведения рыб. Ее образуют особи, близкие по биологическому состоянию и возрасту (при поиске пищи, миграциях, зимовке).
· На стаи образуют не все виды. Стайный образ жизни характерен прежде всего для планктоноядных рыб (например, сельди). Некоторые виды объединяются в стаи только в начальный период жизни, а став половозрелыми, они собираются в стаи только в определенное время (карповые, многие хищники и др.).
· Образование стаи имеет большое приспособительное значение. Стая рыб раньше отдельных особей_ обнаруживает опасность и часто при приближении хищника уплотняется настолько что он не может выхватить из нее отдельных рыб. Хищник вообще быстрее поедает рыб-одиночек. Стая легче избегает орудий лова; на кормовых площадях она скорее обнаруживает скопления пищевых организмов и интенсивнее их поедает. Во время миграций стая лучше ориентируется и быстрее определяет путь.
· Скопление — временная группировка, возникающая из нескольких стай в различные периоды жизни. Миграционные скопления состоят из рыб, совершающих длительные передвижения (миграции), например, к нерестилищам (ход дальневосточных лососей из моря в реки или европейского угря из рек в море), к местам зимовки (ход карповых, сомовых и других рыб осенью в дельту Волги), к местам кормёжки, (скат производителей или молоди с нерестилищ). Нерестовые скопления состоят из половозрелых особей во время нереста (например, скопления сазана на нерестилищах дельты Волги, Дона). Нагульные скопления состоят из рыб, собирающихся на кормовых площадях. Зимовальные скопления состоят из рыб, концентрирующихся на местах зимовки (например, на ямах в устье Волги).
· Стадо (популяция) — локальная самовоспроизводящаяся группировка рыб одного вида разного возраста, которая постоянно обитает в определенном участке водоема. У стада имеются определенные места размножения, нагула и зимовки. Рыбы из разных локальных стад обладают некоторыми морфоэкологическими особенностями.
· Многообразие внутривидовых отношений рыб проявляется в стайном поведении и в пищевых отношениях; в отношениях рыб разного пола, возникающих в период размножения при брачных играх и постройке гнезд; в отношениях родителей и молоди у охраняющих потомство видов, в их влиянии друг на друга через изменение абиотической среды продуктами метаболизма.
· Не менее разнообразны межвидовые связи рыб. У рыб разных видов вырабатываются специфические особенности, помогающие им лучше приспособиться к условиям среды, в которой он формируется. Расхождение спектров питания ослабляет напряженность пищевых отношений. Образование стай, повышение плодовитости, забота о потомстве, выделение яда ядовитыми железами (морской ёрш или морской окунь) или ядовитая икра (у маринки, османа, усача), наличие колючек, шипов облегчают защиту от врагов. Приспособления хищников помогают им в поимке жертв.
· Биотические связи рыб весьма разнообразны. Большое значение в жизни рыб имеют растения. Общеизвестна роль растений в изменении гидрохимического режима водоема: в процессе фотосинтеза на свету из диоксида углерода поглощается углерод и выделяется кислород; массовое развитие и отмирание растений является причиной перегрузки водоема органическими веществами. За счет этого в водоеме происходят значительные колебания содержания кислорода и углекислоты — суточные, сезонные и т. д.
· Растения являются субстратом, на который многие рыбы (фитофильные) откладывают икру. Их заросли служат убежищем от хищников. При этом у многих рыб, постоянно живущих среди растений, вырабатывается специфическая форма тела, делающая их незаметными в этой среде.
· Растения используются в пищу многими рыбами. Низшими растениями питаются многие карповые (белый толстолобик, отчасти плотва и серебристый карась, храмуля, подуст и др.), некоторые цихлиды Tilapia esculenta; высшие растения потребляют белый амур, красноперка, отчасти карась.
· Однако в пресных водах имеются и вредные для рыб растения. Животноядная пузырчатка Utricularia и Aldrovandia захватывают как личинок рыб, так и в большом количестве планктон, вследствие чего обедняется кормовая база в зарослевых участках (в одном растении было насчитано до 3570 шт. захваченных организмов планктона).
· Сине-зеленые водоросли выделяют токсины, которые вызывают увеличение активности фермента, расщепляющего витамин B1 (тиаминаза), вследствие чего у рыб развивается авитаминоз-В1 . Цветение водоема, вызванное массовым развитием этой группы водорослей, кроме резкого ухудшения гидрохимического режима при их разложении может быть причиной отравления рыб, сельскохозяйственных животных и человека.
· Некоторые грибы являются возбудителями различных заболеваний. Широко распространенные грибки из родов сапролегния Saprolegnia и ахлия Achlia поражают и взрослых рыб и икру; грибок бранхиомицес Branchiomyces sp. является возбудителем жаберной гнили, или бронхиомикоза.
· Жизнедеятельность рыб тесно связана с бактериальной флорой. Существует ряд полезных бактерий, некоторые из которых обладают свойством растворять грибы, являющиеся вредителями рыб, другие принимают участие в пищеварении, третьи служат непосредственно пищей молоди рыб и планктонным организмам. С другой стороны, непосредственную опасность для рыб представляют бактерии и вирусы, вызывающие такие болезни, как краснуха карпов, панкреанекроз форелей, фурункулез лососей. Эти заболевания часто приводят к массовой гибели рыб.
· Беспозвоночные играют в жизни рыб огромную роль. Прежде всего велико их пищевое значение. При переходе на питание внешней пищей на ранних личиночных стадиях рыб самые мелкие представители беспозвоночных — простейшие и коловратки — являются основной Пищей молоди. В дальнейшем более крупные объекты — различные представители ракообразных, червей, моллюсков, насекомых — составляют пищевую базу самых различных мирных рыб.
· Вместе с тем представители этих групп могут быть непосредственными врагами молоди рыб. Хищные коловратки, циклопы, водные насекомые — жуки-плавунцы, гладыши, скорпионы, личинки жуков и стрекоз — при массовом развитии могут наносить большой урон молоди в нерестовых прудах. Самое же большое отрицательное значение беспозвоночных заключается в том, что во всех их группах встречаются или непосредственные возбудители, или промежуточные хозяева опасных возбудителей болезней рыб.
· Такие опасные болезни, как хилодонеллез, ихтиофтириоз, триходиниоз вызываются инфузориями хилодонелла Chilodonella sp., ихтиофтириус Ichthyophthirius sp., триходина Trichodina sp.; костиоз — жгутиконосцем костия Costia sp.; вертёж лососевых — споровиком миксозома Myxosoma sp., а черно-пятнистая болезнь передается моллюском катушкой Planorbis sp. Сосальщики паразитируют на жабрах рыб (дактилогирус Dactylogyrus sp. и гиродактилюс Gyrodactylus sp.), в кровеносных сосудах (сангвиникола Sanguinicola sp.), в глазах (диплостома Diplostoma sp.); ленточные черви — гвоздичники Сагуорhyllaeus sp. и ботриоцефалюс Bothryocephalus sp. — поселяются в кишечнике, а круглый червь филометра Phyllometra sp. — в мышцах и т. д. Циклопы являются промежуточными хозяевами ленточного червя ботриоцефалюса, олигохеты — гвоздичника, а моллюск-прудовик диплостомы.
· Связи рыб с позвоночными животными также разносторонни. Земноводные в какой-то степени используются в пищу хищными рыбами. Например, лягушки имеют некоторое значение в питании сома, а головастиков охотно поедают змееголов, форель и щука.
· Лягушки истребляют хищных водных насекомых, чем приносят пользу рыбному населению водоемов. Однако озерные лягушки и тритоны поедают молодь и икру рыб, а урон, наносимый ими промысловым рыбам в местах концентрации молоди, бывает подчас значительным. Кишечники тритонов в нерестовых прудах бывают набиты икрой. В дельте Волги за 2—3 мес. пребывания молоди на полоях лягушки уничтожили на площади 1 га 110 тыс. шт. молоди преимущественно ценных промысловых рыб (сазана, леща, густеры). Головастики могут быть конкурентами в пище молоди рыб, особенно охотно поедая комбикорм в рыбоводных прудах.
· Значение пресмыкающихся ограничивается главным образом их ролью рыбоядов: рыбой питаются водяной уж, черепахи, в меньшей мере крокодилы.
· Водоплавающая птица (птенцы) используется в пищу крупными хищными рыбами (такими, как щука, сом). В местах птичьих базаров благодаря удобрению прибрежных участков моря птичьим пометом значительно увеличивается биомасса пищевых организмов. Использование птичьего помета как удобрения преследует также совместное выращивание рыбы и уток в рыбоводных прудах.
· Птицы, уничтожающие насекомых, являющихся врагами икры и молоди рыб. В свою очередь, рыбоядные птицы — скопа, некоторые орланы, гагары, зимородки буревестники, цапли, бакланы, пеликаны, чайки, утки — требуют для пропитания огромного количества рыбы; например, за один день молодой баклан съедает около 500 г, а взрослый — около 700 г рыбы.
· Многие рыбоядные птицы являются переносчиками опасных заболеваний: чайки — окончательный хозяин лигулы (рис. 47), диплостомы — возбудителя катаракты глаз, чернильной болезни и т. д. Мелкие млекопитающие — мыши-землеройки — могут стать добычей сильных хищных рыб — сома, щуки, ленка, тайменя, хариуса. Сом может захватить даже плывущую собаку. Гроза тропических рек — небольшие рыбки пираньи — мгновенно собираются стаями около раненого животного и, нападая, одолевают его. Они справляются даже с быками.
· Вместе с тем многие млекопитающие относятся к рыбоядным: норки, выдры, водяные крысы, выхухоль, лисы, медведи. Самыми большими потребителями рыбы являются водные млекопитающие — китообразные Ceratea, ластоногие Pinnipedia. Белуха Defphinapterus в огромных количествах поедает лососевых рыб— кету, горбушу, тресковых — сайку, треску, навагу и других рыб. У черноморского дельфина большую долю пищи составляет хамса, шпрот, ставрида, барабулька.
Литература
· Авакян А. Б., Ромашков Е.Г. Рыбы штурмуют плотины. М., 1970.
· Баклашова Т. А. Ихтиология. М., 1980.
· Баранникова И. А. Функциональные основы миграций рыб. Л., 1975.
· Барач Г. Я. Черноморская кумжа. Тбилиси, 1962.
· Бердышев Г. Д. О механизме генетически обусловленной смерти дальневосточных лососей после нереста. — В сб.: Обмен веществ и биохимия рыб. М., 1967, с. 30—37.
· Березина Н. А. Гидробиология. М., 1973.
· Берман Щ. А., Саленице И. К. Пристенное пищеварение у рыб. — Вопр. ихтиологии, 1966, т. 6, № 4, с. 720—724.
· Берген Р. Чувства животных. М., 1972.
· Борисов П. Г., Овсянников Н. С. Определитель промысловых рыб СССР. М., 1954.
· Васнецов В. В. Этапы развития костистых рыб. — В кн.: Очерки по общим вопросам ихтиологии. М., 1953, с. 207—217.
· Веселое Е. А. Определитель пресноводных рыб фауны СССР. М., 1977.
· Винберг Г. Г. Интенсивность обмена и пищевые потребности рыб. Минск, 1956.
· Виноградов В., Воропаев Н., Данченко А. Переселенцы из Америки — сомы-икталурусы. — Рыбоводство и рыболовство, 1974, № 5, с. 10—11.
· Влияние качества производителей на потомство у рыб/Под ред. В. И. Владимирова. Киев, 1965.
· Водоемы Сибири и перспективы их рыбохозяйственного использования/Под ред. Б. Г. Иоганзена. — Материалы совещ. изд-ва Томского ун-та. Томск, 1973.
· Генетика селекции и гибридизация рыб/Отв. ред. Б. И. Черфас. М., 1969.
· Гинзбург А. С. Оплодотворение у рыб и проблема полиспермии. М., 1968.
· Головков Г. За пределами обычного ареала. — Рыбоводство и рыболовство, 1978, № 2, с. 14—15.
· Дмитриев И. Ожившее озеро. — Рыбоводство и рыболовство, 1978, № 4, с. 9—10.
· Ерохина Л,, Виноградов В. Новые объекты рыбоводства и акклиматизации. — Рыбоводство и рыболовство, 1974, №6, с. 12—13.
· Жизнь животных/Под ред. Т. С. Расса. М., 1971, т. 4.
· Зайцев А. В. Строение гипофиза костистых рыб и связь сезонных изменений микроструктуры гипофиза и гонад с условиями размножения. — В кн.: Проблемы современной эмбриологии, 1964, с. 304—308.
· Закономерности роста и созревания рыб/Под, ред. Г. В. Никольского, М., 1971.
· Иванова Н. Т. Материалы к морфологии крови рыб. Ростов н/Д, 1970.
· Исаев А. И., Карпова Е. И. Рыбное хозяйство водохранилищ. М., 1980.
· Карпевич А. Ф. Теория и практика акклиматизации водных организмов. М., 1975.
· Козлов А. Итоги за тридцать лет. — Рыбоводство и рыболовство, 1978, № 2, с. 4—5.
· Константинов А. С. Общая гидробиология. М., 1972.
· Кошелев Б. В, Некоторые закономерности роста и времени наступления первого икрометания у рыб. — В сб.: Закономерности роста и созревания рыб. М., 1971, с. 186—218.
· Краюхин Б. В. Влияние температуры на интенсивность обмена у рыб. — Вопросы ихтиологии, 1961, т. 1, с. 14—19.
· Крыжановский С. Г. Экологические группы рыб и закономерности их развития. — Изв. ТИНРО, 1948, № 27.
· Кудерский Л. А. О путях развития рыбного хозяйства на внутренних водоемах (озера, водохранилища, реки). — Изв. ГосНИОРХ, Л., 1974, т. 87, с. 94—119.
· Кузнецов Б. А. Определитель позвоночных животных фауны СССР. 1974, ч. I.
· Кузьмичев А. Б., Егерева Т. В. Статистические сведения по рыбной промышленности СССР за 1970—1972гг. М., 1973.
· Лавровский В. В. Особенности нереста судака Курского залива как возможного объекта акклиматизации. — Рыбное хозяйство, 1962, № 6, с. 26—30.
· Лапицкий И. И. Направленное формирование ихтиофауны и управление численностью популяций рыб в Цимлянском водохранилище. Волгоград, 1970.
· Лукъяненко В. И. Токсикология рыб. М., 1967.
· Мартышев Ф. Г. Прудовое рыбоводство. М., 1973.
· Матюхин В. А. Биоэнергетика и физиология плавания рыб. Новосибирск, 1973.
· Метелев В. В., Канаве А. И., Дзасохова Н. Г. Водная токсикология, М., 1971.
· Методическое пособие по изучению питания и пищевых отношений рыб в естественных условиях. М., 1974.
· Моисеев П. А., Азизова Н.А., Куранова И. И. Ихтиология. М., 1981.
· Моисеев П. А., Вавилкин А. С., Куранова И. И. Ихтиология и рыбоводство. М., 1975.
· Наумов Н.П., Kapташев Н. Н. Зоология позвоночных. М., 1979.
· Никольский Г. В. Частная ихтиология. М., 1971.
· Никольский Г. В. Экология рыб. М., 1974.
· Никоноров И. В., Шаховский Б. М. Свет ловит рыбу. М., 1967.
· Обмен веществ и биохимия рыб/ Отв. ред. Г. С. Карзинкин. М., 1967.
· Основные особенности поведения и ориентации рыб/Под ред. Б. П. Мантейфеля. М., 1974.
· Панов В. П. Морфологические особенности различных видов карповых рыб в связи с их образом жизни — Докл. ТСХА, 1979.
· Пегель В. А. Физиология пищеварения рыб. Томск, 1950.
· Пенязь В. С., Шевцова Т. М., Нехаева Т. И, Биология рыб водоемов Белорусского Полесья. Минск, 1973.
· Поведение и рецепции рыб/Отв. ред. Г. С. Карзинкин и Г. А. Малютина, М., 1967.
· Поддубный А. Г. Некоторые результаты дистанционных наблюдений за поведением мигрирующих, рыб. — В кн.: Бионика. М., 1965.
· Полтавчук М. А. Биология и разведение днепровского судака в замкнутых водоемах. Киев, 1965.
· Поляков Г. Д. Экологические закономерности популяционной изменчивости рыб. М., 1975.
· Правдин И. Ф. Руководство по изучению рыб. М., 1966.
· Промысловые рыбы СССР/Ред. Л. С. Берг, А. С. Богданов, Н. И. Кожин, Т. С. Расе. М., 1949.
· Протасов В. Р. Зрение и ближняя ориентация рыб. М., 1968.
· Протасов В. Р.,Никольский И. Д. Голоса в мире безмолвия. М., 1969.
· Протасов В. Р. Электрические и акустические поля рыб. М., 1973.
· Пучков Н. В. Физиология рыб. М., 1954.
· Разнокачественность раннего онтогенеза у рыб. /Под ред. В. И. Владимирова. Киев, 1974.
· Рыбохозяйственное освоение внутренних водоемов СССР в 1971 —1975 гг./Шимановская Л. Н., Чистобаева Р. Е., Танасийчук А. Н. и др. — Изв. ГосНИОХР, Л., 1977, т. 126, с. 3—62.
· Рыбы СССР/Лебедев В. Д., Спановская В. Д., Савваитова К. А. и др. М., 1969.
· Смирнов А. И. Биология, размножение и развитие тихоокеанских лососей. М., 1975.
· Смирнова Е. Н. Развитие культурного карпа в зародышевый период. — В сб.: Эколого-морфологические и эколого-физиологические исследования развития рыб. М., 1978.
· Совещание по биологической продуктивности водоемов Сибири. Лимнологический ин-т СО АН СССР/Ред. Б. К. Москаленко, М. Ю. Бекман, К. К. Вотинцев. Иркутск, 1966.
· Соин С. Г. Эколого-морфологические данные о связи каротиноидов с процессами эмбрионального дыхания рыб. — В кн.: Обмен веществ и биохимия рыб. М. 1967.
· Соин С. Г. Приспособительные особенности развития рыб. М., 1968.
· Сорокин С. М. Рыбохозяйственное использование озер СССР. — В сб.: Рыбохозяйственное изучение внутренних водоемов. Л., 1972, № 11, с. 10—23.
· Справочник по болезням рыб/Васильков Г. В., Грищенко Л,И., Енгашев В. Г. и др.; Под ред. В. С. Осетрова. М., 1978.
· Сравнительная физиология животных/Под ред. Т. М. Турпаева. М., 1977.
· Строганов Н. С. Экологическая физиология рыб. М., 1962.
· Суворов Я. Я. Основы ихтиологии. М., 1948.
· Сысоев Н. П. Экономика рыбной промышленности СССР. М., 1977.
· Тамбиев А. X. Летучие вещества, запахи и их биологическое значение. М., 1974.
· Тлеутов Р., Тлеубергенов М. Рыбы Каракалпакии. Нукус, 1974.
· Труды лаборатории основ рыбоводства. Л., 1947, т. I.
· Труды лаборатории основ рыбоводства. Л., 1949, т. II.
· Флерова Г. И. Электрофизиологические исследования обонятельной системы рыб. — В кн.: Сенсорные системы. Морфофизиологические и поведенческие аспекты. Л., 1977, с. 5—22.
· Черфас В. и. Рыбоводство в естественных водоемах. М., 1956.
· Чижик А., Абрамович Л. А. Рыбоводство на юге Украины. Симферополь, 1973.
· Щимановская Л. Н. Состояние рыболовства на внутренних водоемах СССР. — В сб.: Рыбохозяйственное изучение внутренних водоемов. Л., 1972, № 11, с. 3—9.
· Щульман Г. Б. Физиолого-биохимические особенности годовых циклов рыб. М., 1972.
· Balon E. K. Ryby Slovenska. Br., 1966.
· Bauch G. Die einheimischen Susswasserfische. Leipzig, 1966.
· Greenwood P. N. The fisches of Uganda Kampala, 1966.
· Holcik J., Mihalik J. Susswasserfische. Pr., 1968.
· Pflieger W. The fisches of Missuri. Missuri, 1975, p.201— 221.
· Reagan R. E., Pavdue J. B., Eisen E. J. Predicting selection response for growth of chaunel catfish. — J. Heredity, 1976, v. 67, Nl, p. 49—53.