
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Рефераты по косметологии
Рефераты по криминалистике
Рефераты по криминологии
Рефераты по науке и технике
Рефераты по кулинарии
Рефераты по культурологии
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Рефераты по косметологии
Рефераты по криминалистике
Рефераты по криминологии
Рефераты по науке и технике
Рефераты по кулинарии
Рефераты по культурологии
Курсовая работа: Биологические основы выращивания рыбца
Курсовая работа: Биологические основы выращивания рыбца
Содержание
Введение
Глава 1. Биологическая характеристика рыбца в связи со средой обитания и образом жизни
1.1 Общая биология вида
1.2 Эмбриональное развитие
1.3 Постэмбриональное развитие
Глава 2. Влияние различных факторов среды на рыбца
2.1 Влияние температуры
2.2 Влияние освещенности, уровня течения воды на рыбца
2.3 Влияние гидрохимических показателей на рыбца
Глава 3. Управление половыми циклами у рыбца различными методами
Глава 4. Биологические основы кормления рыбца
Глава 5. Транспортировка икры, личинок, молоди и взрослых особей рыбца
Глава 6. Биологические основы акклиматизации
Список используемой литературы
Введение
Рыбное хозяйство в Российской Федерации является комплексным сектором экономики, включающим широкий спектр видов деятельности – от прогнозирования сырьевой базы отрасли до организации торговли рыбной продукцией в стране и за рубежом (Черномашенцев, Мильштейн, 1983).
«Концепция развития рыбного хозяйства Российской Федерации на период до 2020 г.» от 2 сентября 2003г. призвана определить порядок использования биоресурсов Мирового океана российскими рыбаками и внести значительное улучшение в систему управления рыболовством и биоресурсами российских морей. В ней провозглашён переход от свободного и открытого доступа к федеральным биоресурсам – к регулированию добычи через распределение квот методом закрепления промысловых прав.
Принятие концепции совпало со вступлением в действие Соглашения 1995г., с Йоханнесбургским саммитом глав государств (2002г.) и саммитом «восьмёрки» в Эвиане (2003г.). Положение Соглашения и резолюции саммитов требуют существенного улучшения деятельности в области управления биоресурсами и охраны окружающей среды на основе экосистемного и предосторожного подходов.
Положение о Государственном комитете Российской Федерации по рыболовству от 5 сентября 2003г. предполагает, что данный комитет является федеральным органом исполнительной власти, осуществляющим межотраслевую координацию, а также функциональное регулирование в сфере использования, изучения, сохранения и воспроизводства водных биоресурсов и среды их обитания. Госкомрыболовство разрабатывает предложения об общих допустимых уловах, о квотах на вылов водных биоресурсов, разрабатывает и утверждает объёмы товарного выращивания, правила рыболовства, уставы служб органов рыбоохраны, осуществляет государственный учёт и мониторинг водных биоресурсов.
Постановление Правительства Российской Федерации «О квотах на вылов водных биоресурсов» от 10.09.2003г. определяет общий объём квот на вылов водных биоресурсов для обеспечения собственных потребностей; на вылов в промышленных целях во внутренних морских водоёмах, в территориальном море, на континентальном шельфе и в исключительной экономической зоне Российской Федерации в Балтийском, Чёрном, Азовском и Каспийском морях, и в низовьях впадающих в моря рек; на вылов для иностранных государств; на вылов в научно-исследовательских целях, а также в целях акклиматизации и воспроизводства; на вылов для организации любительского и спортивного рыболовства.
Целью развития рыбного хозяйства в Российской Федерации является достижение устойчивого функционирования рыбохозяйственного комплекса на основе сохранения, воспроизводства и рационального использования водных биологических ресурсов, развития аква – и марикультуры, обеспечивающего удовлетворение внутреннего спроса на рыбную продукцию, продовольственную независимость страны, социально-экономическое развитие регионов, экономика которых зависит от прибрежного рыбного промысла. При этом должны быть созданы условия для повышения эффективности экспорта рыбной продукции и ее конкурентоспособности и оптимизации структуры управления рыбохозяйственным комплексом (Моисеев, 1981).
По данным А. К. Никонорова и В. И. Ананьева (2007), достижение этой цели возможно при формировании комплексного подхода к государственному управлению развитием рыбного хозяйства в Российской Федерации. Создание эффективной системы управления, предусматривает решение следующих задач:
– разработка нормативной правовой базы в области рыбного хозяйства, соответствующей задачам его эффективного развития;
– формирование и реализация механизма долгосрочного и эффективного управления водными биологическими ресурсами, обеспечивающего прозрачность системы их распределения;
– сохранение и рациональное использование водных биологических ресурсов, уменьшение промысловой нагрузки на эти ресурсы;
– развитие искусственного воспроизводства водных биологических ресурсов, формирование генофондовых коллекций и маточных стад ценных видов этих ресурсов;
– разработка государственной социальной стратегии в области рыбохозяйственного комплекса, обеспечивающей оптимальную занятость и доходы населения в субъектах Российской Федерации, территории которых прилегают к морскому побережью.
В мировом рыбном хозяйстве производство продукции аквакультуры за последние четыре десятилетия выросло более чем в 100 раз и в 2004 г. превысило 50 млн т, что составляет более 55 % общемирового вылова рыбы и других гидробионтов. . В конце прошлого века каждая четвертая съедаемая в мире рыба .была выращена при непосредственном участии человека. В последние годы аквакультура во многих странах превратилась в одну из наиболее быстропрогрессирующих отраслей производства продовольствия.
Рыбохозяйственный фонд внутренних пресноводных водоемов России включает 22,5 млн га озер, 4,3 млн га водохранилищ, 0,96 млн га сельскохозяйственных водоемов комплексного назначения, 142,9 тыс. га прудов и 523 тыс. км рек. Темпы роста производства продукции аквакультуры значительно превышают таковые в рыболовстве, что подтверждает общемировые тенденции ускоренного развития искусственного разведения гидробионтов против упрощенного изъятия естественных биологических ресурсов Мирового океана. В то же время, несмотря на то, что объемы производства продукции аквакультуры возрастают, ее доля в объеме ВВП страны снизилась с 0,16 % в 2000 г. до 0,09 % в 2004 г (Печников, Шурухин, 2006).
Важнейшим определяющим фактором развития аквакультуры на ближайшую перспективу является повышение доли продукции аквакультуры в продовольственном обеспечении населения и улучшение качества питания за счет увеличения рыбной продукции в продовольственном рационе. Исходя из этого, рассчитаны необходимые объемы производства продукции аквакультуры на период до 2015 г.
Объектом исследования является рыбец – очень ценная промысловая рыба. По данным ВНИРО вылов рыбца в 2001 и 2002 годах в водоёмах РФ составил 1760 т. и 1897т. соответственно, что составляет чуть больше 1% от общего вылова карповых рыб. Незначительность промыслового использования рыбца связано с повсеместным снижением его численности, вызванное зарегулированием стока рек, их загрязнением, всё возрастающим водопотреблением и нерациональным выловом. По мнению Л. П. Астанина, 1968 г. для сохранения вида и рационального его использования необходимо введение полного запрета на лов рыбца на нерестилищах, в течение всего срока нереста и в период нерестовых миграций в реки.
Целью работы является глубокое изучение биологических особенностей рыбца, так как на основе этих знаний разрабатываются правила рыболовства, включающие допустимый объём улова, ограничения на лов неполовозрелой и мелкой рыбы и другие меры направленные на рациональное и эффективное использование ресурсов. Это даёт нам возможность получения высоких устойчивых уловов при сохранении на должном уровне воспроизводительной способности эксплуатируемой популяции рыбца.
Глава 1. Биологическая характеристика рыбца в связи со средой его обитания и образом жизни
1.1 Общая биология вида
Надкласс: Бесчелюстные (Gnathostomata).
Ряд: Рыбы (Pisces).
Класс: Костные рыбы (Osteichthyes).
Подкласс: Лучепёрые (Actinopterygii).
Надотряд: Костистые рыбы (Teleostei).
Отряд: Карпообразные (Cypriniformes).
Подотряд: Карповидные (Cyprinoidei).
Семейство: Карповые (Cyprinidae).
Род: Рыбец (Vimba).
Вид: Рыбец (Vimba Vimba)
Рыбец (рис. 1.) — широко распространённый вид, численность которого повсеместно снижается. Включён в список редких и исчезающих рыб Европы и в Красную книгу Белоруссии.
Тело относительно высокое. Рыло выдаётся вперёд. Рот нижний, полулунной формы. Между затылком и началом спинного плавника имеется бороздка, свободная от чешуи. Позади брюшных плавников киль, не покрытый чешуёй. В спинном плавнике 3 не ветвистых и 8-9 ветвистых лучей, в анальном 3 не ветвистых и 16-22 ветвистых лучей. В боковой линии 49-63 чешуи. Имеются глоточные зубы однорядные, обычно 5-5.Жаберные тычинки короткие, редкие (16-20). Спина голубовато-серая , брюхо серебристо-серое, грудные, брюшные и анальные плавники бледно-желтоватые. Ко времени нереста рыбец приобретает брачный наряд наиболее ярко выраженный у самцов: спина становится угольно-чёрной; брюхо, а также брюшные и анальный плавники краснеют. На теле и голове у самцов появляются белые эпителиальные бугорки (Решетников, 2002).
Вид близкородственный к лещам, отличающийся от них более коротким анальным плавником (15-22 ветвистых луча), наличием за спинным плавником ясно выраженного спинного киля, покрытого чешуей; нижним ртом. Сыртью этот вид называют на западе, рыбцом – на юге Европы (Рыжов, 1987).

Рис. 1. Рыбец
Встречается в бассейнах Северного, Балтийского, Чёрного, Азовского и Каспийского морей. На юг распространён до Эгейского и Мраморного морей- реки Вардар, Марица, Струма, места на Балканском полуострове и реки Западной Турции (Астанин, 1968; Рыжов, 1987; Решетников, 2002). В пределах столь широкого ареала образует ряд подвидов, из которых основными являются три:
1.Северная сырть Vimba Vimba Vimba- бассейны Северного и Балтийского морей (реки Эльба, Везер, Висла, Нямунас, Даугава, Финляндии и южной Швеции, а также прибрежная полоса балтийского моря). Достигает длины 50 см и массы 3 кг. Продолжительность жизни 11 лет.
2.Черноморская сырть Vimba Vimba carinata- бассейны Чёрного и Азовского морей (реки Дунай, Днестр, Южный Буг, Днепр, Кубань, Дон и участки этих морей). Длина до 40 см и масса до 1 кг. Продолжительность жизни 8 лет.
3.Каспийский рыбец Vimba Vimba persa- бассейн Каспийского моря (преимущественно реки западного и южного побережий- Кура, Терек и прибрежная часть моря). Длина до 30 см и масса до 400 г. Продолжительность жизни 7 лет.
Хорошо прижился в ряде водохранилищ - Каунасском на Нямунасе, Цимлянском на Дону, Сенгилеевском на Кубани, Ткибульском на Риони и др.
Проходная и полупроходная стайная рыба, поднимающаяся в реки для размножения. Идёт дважды в году: весной и осенью. Рыбцы осеннего хода зимуют в пресной воде. Рыбцы, обитающие в водохранилищах, идут метать икру во впадающие реки. Половозрелыми сырть и черноморский рыбец становятся в возрасте 4-5 лет, а каспийский рыбец в 3-4 года. На нерест сырть и рыбец поднимаются в реки и мечут икру на перекатах с каменистым, обычно галечным грунтом. Глубина на нерестилищах 0,5-1 м, скорость течения 0,7-0,8 м/с. В юго-западной части Балтийского моря и дельте Терека сырть может нереститься на растительности в небольших заливах и в устье рек. Рыбец Днепровско-Бугского лимана для размножения мигрирует главным образом в Днепр и его рукава. Лучшими нерестилищами для рыбца являются россыпи мелкого камня и ракушечника, образовавшиеся при размыве берегов и заливаемые водой только в период половодья. Эти участки значительную часть лета бывают вне воды, хорошо обсыхают, проветриваются, поэтому на них мало обрастаний и они менее заиленные. Небольшая глубина, относительная прозрачность воды позволяют наблюдать за нерестом с берега или, еще лучше, с лодки. Можно видеть, что готовые или близкие к нересту рыбцы заходят на нерестилище одновременно небольшими группами. Нерест растянут с мая по июль. На севере ареала он начинается позднее, чем на юге. Икра у самок созревает 2-4 порциями. Температурный диапазон, при котором происходит размножение, очень широк- от 12 до 24 С. Самку обычно сопровождают 5-7 самцов. Самки быстро передвигаются, выбирая места, подходящие для откладки икры. В момент икрометания самка становится против течения. Ее голова наклонена вниз, а хвостовой стебель поднят несколько вверх. Двигая хвостом, она медленно продвигается вперед и при этом выпускает икру. Находящиеся по бокам самцы поливают икру молоками. Нерест рыбца наиболее активно протекает в утренние (с 5 до 12) и вечерние (с 16 до 24) часы. С наступлением ночи нерест прекращается, рыбец отходит от берегов. На нерестилищах самцов гораздо больше, нежели самок. Во время нереста рыбца на тех же местах держатся стайки уклей, пескарей, бычков. Судя по наблюдениям, они поедают только ту икру, которая задерживается на верхней стороне камней. Обычно икра заносится под камни. Часто на стороне нерестилища, обращенной к течению, икра размещается плотным слоем, наподобие дерна. Подобные мощные кладки икры образуются благодаря значительным концентрациям производителей, а также потому, что икра от нескольких самок заносится в одни и те же ямки и щели. Скопления икры лежат рыхло и хорошо промываются водой. Икра выметывается и созревает порциями, чаще наблюдается 3 порции икры, но у самок, нерестующих первый раз, бывает 2 порции. Возможно, что у старых самок число порций икры также сокращается. Так, у самок восьмилетнего возраста и старше можно обнаружить только одну порцию икры или они и вовсе пропускают нерест. В р. Дон рыбец заходит в течение круглого года, особенно заметен его ход с осени до поздней весны, но основной ход весной (Рыжов, 1987).
Развитие икры при температуре 19-21 оС продолжается 2-2,5 суток. Выклюнувшиеся личинки первое время лежат неподвижно между камнями. Плодовитость в зависимости от размера самок колеблется в очень широких пределах- от 11,4 до 136,4 тыс. икринок. После нереста производители быстро скатываются в море. Молодь может задерживаться в нерестовых реках на продолжительное время- от 4-5 мес. до 2-2,5 лет. В море рыбец держится вблизи берега, на глубине не более 25 м. Питается он здесь мелкими моллюсками, личинками насекомых, червями, бокоплавами. Молодь в пресной воде поедает зоопланктон.
В настоящее время условия естественного нереста для сырти в бассейне Балтийского моря, для рыбца в реках Кубань, Дон, Днепр и Днестр осложнились. В результате строительства плотин часть нерестилищ попала в зону затопления, а гидрологический режим на нерестилищах, оставшихся в нижнем бьефе, резко изменился. Так, в условиях нижнего бьефа днестровский рыбец выметывает первую порцию икры, а вторая порция у многих особей не развивается, а подвергается перерождению и рассасывается. В некоторых водохранилищах (например, Дубоссаровском) создаются свои стада рыбца, они находят благоприятные условия для нереста в верхнем бьефе. Но численность такого рыбца ограничена кормовыми ресурсами водохранилища. Для поддержания запасов проходного рыбца создаются искусственные нерестилища в нижнем бьефе. Уже давно рыбаки для привлечения рыбца в нерестовую пору специально насыпают участки гравия, так называемые гребли. Исследования последних лет показали, что нерест на греблях проходит вполне успешно. Кроме того, икру рыбца инкубируют и на рыбоводных заводах, а молодь подращивают. Сырть и рыбец - ценные промысловые рыбы (Решетников, 2002).
1.2 Эмбриональное развитие
Рыбец откладывает икру на галечном грунте на перекатах в местах с быстрым течением и благоприятным кислородным режимом при температуре обычно 17 °С и выше. Его развитие в раннем периоде онтогенеза проходит в этих условиях и приспособлено к ним.
Икра обычно желтого цвета, но встречаются икринки с зеленоватым оттенком, бесцветные и др. Средний диаметр икры 1,5-1,8 мм с небольшим перивителлиновым пространством, она полиплазматическая. По количеству цитоплазмы занимает одно из первых мест среди икры рыб семейства карповые. Диаметр желточного мешка в среднем 1,2 мм. Оболочка икры клейкая. Продолжительность развития икры рыбца до выхода из оболочек эмбрионов зависит прежде всего от температурных условий. Однако для развития икры и выклева необходимо, как установлено, определённое количество тепла.
Эмбриональный период развития рыбца состоит из девяти этапов.
На первом этапе происходит образование перивителлинового пространства и бластодиска (рис. 1а, б). У неоплодотворенной икринки (см. рис. 1,а) оболочка плотно прилегает к желтку. Началом первого этапа онтогенеза является образование зиготы. Этап продолжается до начала дробления. Через несколько минут после оплодотворения в икре, находящейся в воде, происходят изменения, связанные с проникновением воды в икринку. Это приводит к отслоению оболочки от желтка, образованию перивителлинового пространства. Процесс набухания икры при температуре 17 - 19 °С длится 1,25 часа. Диаметр икры увеличивается в среднем на одну треть. Одновременно в период набухания образуется зародышевый диск, или бластодиск (рис. 2,б).
Активация икринок, вызванная оплодотворением, приводит глубоким изменениям обмена веществ. В течение первого часа после оплодотворения, когда наступает резкое оводнение икринок, относительное содержание сухих веществ снижается с 30-32 до 10-12 % и примерно в таком количестве остается до выклева эмбриона. Содержание гликогена - основного источника энергии в период образования бластодиска - уменьшается в 2 раза, а величина аденозинтрифосфорной кислоты (АТФ), занимающей центральное место в энергетическомобмене, снижается почти в 3 раза.
На втором этапе происходит дробление бластодиска от появления первой борозды до морулы мелких клеток. Икринка проходит ряд стадий развития. В возрасте трёх часов наступает стадия дробления, появляется первая борозда, делящая бластодиск на две клетки-бластомера (рис. 2,в), а затем наступают стадии четырёх (рис.2,г), восьми (рис. 2,д) бластомеров и т. д. Через 6 часом от момента оплодотворения наступает стадия морулы крупных клеток (рис. 2,е). Далее клетки бластодисков всё больше дробятся. Наступает стадия морулы мелких клеток. Этап длится около 9 часов.
Третий этап – образование бластулы. Длится 6 часов. Процесс дробления прекращается. Между бластодиском и желтком возникает небольшая полость, или бластоцель, и образуется стадия бластулы (рис. 2,ж). Бластула – это своеобразное многоклеточное образование - бластодерма, расположенная на анимальном полюсе желтка. Края бластодиска наползают на желточный мешок. Анимальный участок желточного мешка вдавливается в эту полость, отчасти заполняя её.
В целом процесс дробления сопровождается значительными внутренними энергетическими затратами. За этот период показатель АТФ снижается почти в два раза.
В рыбоводной практике на стадиях 4-8 бластомеров второго этапа дают оценку качества икры по нормальному дроблению. Образование разномерных, асимметрично расположенных бластомеров свидетельствует об аномальном развитии икры. Именно на стадиях дробления от 4-8 бластомеров до ранней морулы определяют и процент оплодотворения икры.
На четвёртом этапе происходит образование гаструлы. Гаструляция начинается с обрастания желтка многослойной бластодермой (рис. 2,з), которая подворачивается и образует в одном из участков краевой зоны зародышевый узелок, и даёт начало образованию зародышевых пластов. Его хорошо видно на стадии замыкания желточной пробки (рис. 2,и). У тела зародыша заметен расширенный головной отдел. Гаструляция завершается полным обрастанием бластодермой всего желтка. Желточный мешок вытягивается, принимая удлинённую, шаровидную форму. Этап длится 7 часов.
Во время гаструляции происходит существенная структурная перестройка, в результате которой образуются три зародышевых листка: эктодерма, мезодерма и энтодерма.
Обмен веществ во время гаструляции имеет особенности. В этот период создаются основы органогенеза. После гаструляции количество фосфора АТФ и небелкового азота снижается, а количество общего белка увеличивается. Процесс гаструляции является наиболее уязвимым к воздействию факторов внешней среды. Гаструляция всегда сопровождается повышенной гибелью икры. Поэтому учет отхода целесообразно проводить после прохождения этой стадии, а не раньше.
На пятом этапе происходит дифференциация головного и туловищного отделов зародыша. Наблюдается утолщение головной и хвостовой частей зародыша. Происходит также дифференцировка зачатков основных органов. В течение этапа формируется 21 – 24 туловищных миотома (рис. 2,к). Этап длится 17 часов.
На шестом этапе образуются глазные бокалы, хрусталики при продолжающейся сегментации тела (рис. 2,л). За глазными пузырями в области продолговатого мозга появляются слуховые плакоды (рис. 2,м) Обособляется хвостовой отдел и зародыш начинает двигаться. В результате обособления хвостового отдела и роста в длину зачатка кишечной трубки желток приобретает грушевидную форму. В глазах отчетливо виден хрусталик (рис. 2,н). Тело эмбриона совершает слабые движения. Наблюдается сегментация хвостового отдела. К этому времени сегментация тела почти заканчивается. В глазах появляется черный пигмент. Различаются отделы головного мозга. В слуховых капсулах образовываются оттолиты (рис. 2,о). При обособлении хвостового отдела и пигментации глаз наступают определенные изменения в обмене веществ: показатель АТФ вновь возрастает до исходной величины, однако содержание белка и небелкового азота остается прежним, как при гаструляции. Этап длится 11 часов.
На седьмом этапе у зародыша заканчивается сегментация хвостового отдела и начинается распрямление головы, в связи с её обособлением от желточного мешка. Рот зачаточный в виде ямки. Появляются форменные элементы в крови, развивается эмбриональная сосудистая система, несущая дыхательную функцию, в виде кювьеровых протоков на передней расширенной части желточного мешка и нижней хвостовой вены в анальном отделе общей непарной плавниковой складки. Кровь по мандибулярным дугам аорты из сердца поступает в голову, а в туловище – по спинной аорте, которая, загибаясь в хвосте, переходит в хвостовую вену, спускающуюся в анальный отдел общей непарной плавниковой складки. Грудные плавники зачатковые. Длительность этапа 58 – 72 часа.
Эта стадия зародыша рыбца, как и других рыб, наиболее подходит для перевозки икры в условиях изотермических ящиков, где возможно некоторое охлаждение, способствующее замедлению развития.
На восьмом этапе из оболочки вылупляется эмбрион. Голова окончательно распрямляется и полностью отделяется от желточного мешка. Вначале этапа желточный мешок грушевидный, к концу его становится сигаровидным. Рот нижний, неподвижный, открытый. В глазах увеличивается количество малинового пигмента, они становятся чёрными, к концу этапа в них появляется гуанин(рис. 2,о.). В слуховых пузырьках дифференцируются полукружные каналы. Формируются жаберно-челюстной аппарат, печень, желточный и плавательный пузыри. Кишечник имеет прямую сдавленную трубку без просвета (рис. 2,р). Кювьеровы протоки укорачиваются. Сосудистая сеть нижней хвостовой вены достигает максимального развития. В общей непарной плавниковой складке формируются хвостовая, спинная и анальная лопасти. Функционируют железы приклеивания. На теле появляется пигмент, в кровяных тельцах – гемоглобин. Предличинки имеют отрицательную реакцию на свет.Они приклеиваются к субстрату,В конце этапа предличинки начинают время от времени всплывать .Длительность этапа 79 часов.
После выхода эмбриона из оболочки существенные изменения происходят и в обмене веществ. Если гликоген является основным источником энергии зародыша, то главным в эндогенном питании предличинки является жир. Его запасы в два раза выше (2-2,5 %), чем гликогена (0,7—1,2 %). Меняются и другие показатели обмена. Содержание белка увеличивается до 11-13 %, сухих веществ - до 19-20 %, фосфора - до 300-360 %.
На девятом этапе рот становится подвижным. Увеличиваются слуховые пузыри и оттолиты. Приобретают подвижность грудные плавники. Наряду с эмбриональными органами дыхания, функцию дыхания выполняют жабры. В начале этапа начинается кровообращение в лепестках жаберных дуг. К концу этапа лепестки удлиняются и становятся разветвлёнными. Кювьеровы протоки уже не функционируют как эмбриональный орган дыхания. Значительно уменьшается объём желточного мешка. Предличинки постепенно утрачивают отрицательную реакцию на свет. Они начинают всплывать на поверхность воды и захватывать пузырьки воздуха, наполняя им плавательный пузырь, а наполнив, переходят к постоянному плаванию в толще воды. Этап длится 48 часов, личинка достигает 8,0 – 8,6 мм.
Необходимо обратить внимание на очень важное обстоятельство, которое надо учитывать в рыбохозяйственной практике и особенно в современном рыбоводстве при широком использовании заводского способа получения личинок рыбца – икра рыб в процессе эмбрионального развития проходит ряд критических периодов, когда наблюдается повышенная чувствительность эмбрионов к различным абиотическим факторам среды (температуре, газовому составу воды, солености, механическому воздействию и др.). Это связано с тем, что в критические периоды происходят значительные изменения в перестройке обмена веществ развивающегося зародыша.
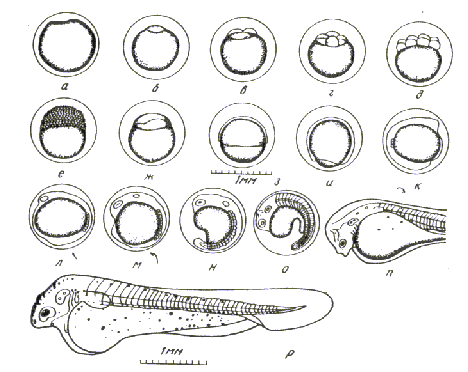
Рис.2. Эмбриональный период развития икры рыбца
Критическими периодами в развитии икры рыбца, как у большинства нерестящихся весной рыб, являются следующие стадии: начало дробления до морулы мелких клеток, гаструляция, стадия перед выклевом и в период выхода зародыша из оболочки. Именно на этих стадиях эмбриогенеза, особенно в начале дробления, вступления икры в стадию ранней гаструлы и замыкания желточной пробки, перед вылуплением и в момент выхода эмбриона из оболочки, наблюдается повышенная гибель зародышей. После прохождения критического периода гибель эмбрионов наблюдается не сразу, а спустя некоторое время, чаще перед наступлением следующей стадии развития.
В момент критических периодов необходимо особенно стремиться к созданию оптимальных условий для развития икры: поддерживать в инкубационных аппаратах постоянный и повышенный расход воды, недопускать резких температурных перепадов, оберегать икру от различных механических воздействий и т. д.
1.3 Постэмбриональное развитие
Постэмриональное развитие включает личиночный и мальковый периоды. В раннем периоде с момента вылупления из оболочки рыбец проходит 7 этапов развития..
Личиночный период.
Этап первый (В)- личинка достигает длины 8,6-9,3 мм , длится 2 суток. На этом этапе для личинок характерен смешанный тип питания: личинки захватывают пищу извне (инфузории, коловратки, одноклеточные водоросли) и питаются ещё за счёт содержимого желточного мешка. Рот не совсем конечный, не полностью закрывающийся. Дыхательная функция выполняется главным образом жабрами. Роль эмбриональных дыхательных систем уменьшается. Сеть сигментных сосудов в значительной степени редуцируется. Плавательный пузырь однокамерный, заполнен воздухом. Желточный мешок имеет перетяжку. Дегенерируют железы приклеивания. В течение этапа формируется до 4 лепидотрихий (рис. 3).
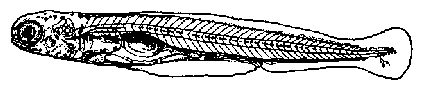
Рис. 3. Этап В развития рыбца
Этап второй (С1) –личинка размером 8,9-9,7 мм. Длительность этапа 4 суток. Желточный мешок полностью рассасывается и личинка переходит на экзогенное питание. Рот конечный. Закладываются хрящевые нижние дужки позвонков, продолжается формирование липидотрихий (рис. 4.). Питаются одноклеточными водорослями, коловратками, инфузориями, молодью кладоцер и копепод.
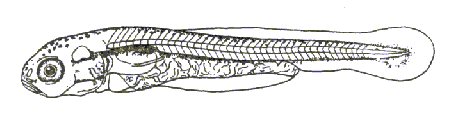
Рис. 4. Этап С1 развития рыбца
Этап третий (С2 )– длина личинки 9,3 - 11,7 мм. Продолжительность этапа 8 суток. Рыло значительно удлиняется. Жаберные крышки начинают окостеневать, но они ещё не достигают пояса грудных плавников. Исчезают эмбриональные дыхательные системы. Редуцируются выросты сегментных сосудов. Нижняя хвостовая вена частью спрятана под миотомами. Плавательный пузырь однокамерный, но с зачатком передней камеры. Кишечник в виде трубки. Конец хорды загнут под тупым концом вверх, под ним имеются разросшиеся гипуралии и отходящие от них косо вниз лепидотрихии (рис. 5.). В течение этапа формируется полное число лепидотрихий (до 20).питаются одноклеточными водорослямии молодью кладоцер и копепод.
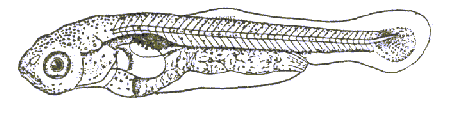
Рис. 5. Этап С2 развития рыбца
Этап четвёртый (D1)-длина личинок 11,3-13,9 мм. Продолжительность этапа 6 суток. Плавательный пузырь становится двух камерным. Передняя камера его наполняется воздухом. В хвостовом плавнике костные лучи. В спинном и анальном плавниках мезенхимные лучи. (рис. 6). Питаются, как и на предыдущем этапе.
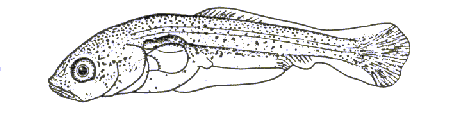
Рис. 6. Этап D1 развития рыбца
Этап пятый (D2) – длина личинок 12,3-15,7 мм. Длительность этапа 6 суток. Края жаберных крышек заходят за пояс грудных плавников. Строение спинного, анального и хвостового плавников становится таким же, как у взрослых рыб. В этих плавниках развиты костные лучи. Грудные и брюшные плавники увеличиваются, в них появляются мезенхимные лучи. (рис. 7). В основном питается взрослыми формами кладоцер и копепод, личинками фитофильных хирономид.

Рис. 7. Этап D2 развития рыбца
Этап шестой (Е) – личинки длиной 15,7-18,8 мм. Длительность 13 суток. Жаберные крышки полностью окостенели. Появились первые две петли кишечника. Во всех парных и непарных плавниках хорошо развиты костные лучи. Редуцированы остатки плавниковой складки позади спинного и анального плавников. Брюшные плавники выступают за края преанальной складки. Спектр питания тот же, что и на пятом этапе. (рис. 8).

Рис. 8. Рыбец на этапе Е
Мальковый период.
Этап F – личинка достигает длины 16,0-19,0 мм. Продолжительность этапа 6 суток. Рот перемещается в нижнее положение. Обонятельное отверстие полностью разделено перегородкой на два. Мягкие лучи в непарных плавниках становятся ветвистыми. Остатки плавниковой складки полностью редуцируются. Заканчивается формирование позвонков и рёбер. Вентральные концы миотома смыкаются по средней линии. Жаберные крышки и бока приобретают серебристую окраску. В конце этапа на теле появляется чешуя. Питаются зоопланктоном и зообентосом. (рис. 9).

Рис. 9. Этап F развития рыбца
В рыбоводной практике сроки пересадки молоди в мальковые или выростные пруды определяются не возрастом, а этапом развития молоди. Зарыбление мальковых или выростных прудов рекомендуется осуществлять личинками на этапе смешанного питания. Продолжительность подращивания определяется временем, необходимым для завершения личиночного периода развития (Астанин, 1968).
Продолжительность каждого этапа зависит от температуры воды, обеспеченности пищей, гидрохимических условий и селекционных особенностей данного объекта.
Глава 2. Влияние различных факторов среды на рыбца
Рыбы, как и все другие организмы, обитающие в водоемах, находятся в тесном взаимодействии с абиотическими факторами среды — совокупностью условий неживой природы, и биотическими факторами среды, включающими в себя разнообразные отношения между организмами. Грунт и вода с ее химическим составом и физическими показателями являются абиотическими факторами среды, действующими в водоемах. Биотические взаимоотношения подразделяются на межвидовые и внутривидовые.
Внешняя среда влияет на все жизненные процессы, происходящие в организме рыбы: дыхание, питание, кроветворение и кровообращение, нервную деятельность, размножение, рост и развитие. Рыба на разных стадиях своего развития и в различные периоды жизни неодинаково реагирует на условия внешней среды (Берлянд, 1953).
Чтобы правильно оценить ту огромную роль, которую играют факторы внешней среды в жизни рыб, достаточно рассмотреть лишь наиболее важные из них, с которыми чаще всего приходится сталкиваться специалисту, работающему в области рыбоводства: температуру воды, освещенность, уровень и течение воды, гидрохимический режим.
2.1 Влияние температуры воды
Температура воды является одним из факторов, оказывающих большое воздействие на отправление жизненных функций рыбы, определяющих ее рост и развитие. Этот фактор действует на рыбу как непосредственно — изменяя интенсивность ферментативных процессов, происходящих в организме, активность потребления пищи, характер обмена веществ, ход развития половых желез и прочее, так и косвенно, оказывая первое влияние на улучшение или ухудшение развития естественной кормовой базы. Температура является также внешним стимулом, определяющим для физиологически подготовленного организма рыб начало миграций, нереста и зимовки. Температурные условия, при которых все жизненные процессы протекают в организме нормально, принято называть оптимальными. Исходя из оптимальных температурных условий, все виды рыб условно подразделяют на теплолюбивых и холодолюбивых. (Иванов, 1988)
Теплолюбивые рыбы, к которым относится объект вселения – рыбец может жить в водоемах, в которых температура воды изменяется в течение года от 0 до 30 °С и даже несколько выше. Температурный диапазон размножения рыбца 12 - 24 оС. Икра развивается при температуре 19 – 21оС (Рыжов, 1987).
При постепенном повышении или понижении температуры (по отношению к оптимальным температурным условиям) нормальные течения жизненных процессов в организме рыбы нарушаются. С повышением температуры активизируются многие жизненные процессы: увеличиваются частота сердцебиений и двигательная активность, потребление кислорода и усвояемость корма, усиливается обмен веществ, рыба лучше растет. Температура влияет на время и продолжительность созревания половых продуктов у рыб, начало и конец нереста, длительность инкубационного периода икры и т. д.
При повышении температуры воды на каждые 10 °С частота сердцебиений у рыб увеличивается в 1,8 – 2,5 раза. C повышением температуры воды увеличивается и потребление кислорода рыбой.
Переваримость корма, а, следовательно, и усвояемость его также увеличиваются с повышением температуры. При температуре 0 °С обмен веществ, примерно в 6 раз ниже, а при 30° С в 2 раза выше, чем при 20 °С.
Резкое изменение температуры воды, если даже она и не выходит за границы оптимальных температурных условий, вызывает у рыб нервный шок, который приводит обычно к гибели (Иванов, 1988).
Изменения температуры воды оказывают влияние на некоторые миграции рыб. Однако, отмечая влияние температуры воды на миграции рыб, необходимо указать, что она при этом не является единственным и определяющим фактором, а воздействует в совокупности с другими условиями среды (уровень и скорость течения воды в реках, прозрачность воды, продолжительность светового дня и др.).
В зависимости от температуры воды изменяется количество растворенного в воде кислорода, который необходим для дыхания рыб. Так, при понижении температуры содержание кислорода в воде повышается, а при повышении снижается (Касымов, 1987).
Температура воды оказывает также большое влияние на развитие в водоеме кормовых организмов, являющихся пищей для рыб. Отклонение температуры воды от оптимальной для организмов зоопланктона и бентоса вызывает снижение их численности и биомассы.
Температурный режим может оказывать косвенное влияние и на анатомическое строение рыб: количество позвонков и лучей в спинном и анальном плавниках (Карпевич, 1981).
Исходя из изложенного выше, можно сказать, что температура воды имеет огромное значение в жизни рыб. Поэтому рыбовод должен постоянно контролировать температуру воды на рыбоводных предприятиях, а при необходимости, обусловленной биологической целесообразностью, изменять ее до желаемой величины.
2.2 Влияние освещённости, уровня и течения воды на рыбца
Течения влияют на физические, химические и биологические процессы, происходящие в водоемах. Теплые течения, приносящие тепло в холодноводные районы, создают благоприятные условия для развития кормовых организмов, а следовательно, и для рыб (Карпевич, 1981). В местах соприкосновения теплых и холодных течений образуются фронтальные зоны, в пределах которых происходят интенсивное вертикальное перемешивание водных масс и обогащение их биогенными элементами, интенсивное развитие фито- и зоопланктона, а следовательно, создаются условия, благоприятные для рыб.
Течения играют решающую роль в ориентации рыб в водном пространстве, обуславливают так называемые хоуминговый и стрейнинговый эффекты. Также они имеют большое значение для распространения пелагической икры и личинок у многих видов рыб (тунцы, речной угорь и др.) (Павлов, 1979).
Установлено большое значение круговых течений в различных областях океана для развития пелагической икры и личинок рыб. В пределах локальных круговых течений икра и личинки многих промысловых рыб развиваются до стадии активно двигающегося малька. При нарушении круговых течений они прямолинейными течениями уносятся далеко от районов нереста, нередко попадая в районы с неблагоприятным гидрологическим режимом и погибая (Черномашенцев, Мильштейн, 1983).
В реках движущийся поток оказывает влияние на строение рыб. Одни из них имеют вальковатое тело и являются хорошими пловцами, как например рыбец. Это связано с тем, что рыбец является проходной рыбой и для достижения мест нереста ему приходится преодолевать большие расстояния, двигаясь вверх по течению. У других рыб тело удлиненное, позволяющее им жить между камнями (пескарь, голец) (Павлов, 1979).
Для большинства видов проходных рыб, в том числе и рыбца, течение воды в реке является стимулирующим фактором для созревания половых продуктов и при нересте. Вместе с тем уровень воды в реке также имеет большое значение для этих рыб. Поступление в море паводковых вод, повышающих уровень воды в реке и скорость ее течения, является наряду с температурой внешним раздражителем нерестовой миграции рыб. Весенний максимум нерестового хода этих рыб приурочен к резкому подъему уровня воды в реках и большой ее мутности. В это время производители заходят в реки и продвигаются к нерестилищам, где они размножаются при наступлении нерестовых температур. Однако их нерест происходит только в том случае, если на нерестилищах имеются галечный грунт и определенные скорости течения. При отсутствии этих условий половые клетки производителей резорбируются (Астанин, Саманеева, 1968).
Свет. Основным источником света в воде является солнечная радиация. Солнечный свет поглощается поверхностным слоем воды, и только 0,45% его достигает глубины 100 м. Прозрачность воды, значительно влияющая на освещённость, зависит от содержания в толще воды неорганических и органических взвешенных частиц, а также мельчайших растительных и животных организмов. Большое количество взвешенных частиц глины и песка в воде вызывает отмирание фито- и зоопланктона, затрудняет дыхание рыб и ухудшает их питание. Мутность воды, образованная взвесью частиц из отмерших растительных и животных организмов, ухудшает гидрохимический режим водоема (Моисеев, 1981).
Освещение водной среды отличается от освещения воздушной. Волны света разной длины достигают разных глубин. Инфракрасные (тепловые лучи) поглощаются в самом верхнем (до 1 м) слое воды. На глубину 5 м проникает лишь 10% красных лучей, на глубину 13 м — лишь 10 % зеленых лучей, а на глубины 500 м и более проникают только фиолетовые и ультрафиолетовые лучи.
В связи с такой освещенностью водной среды глаз рыб в отличие от глаза человека менее чувствителен к красным лучам и более чувствителен к желтым, зеленым, синим и фиолетовым. Рыбы чувствительны и к ультрафиолетовым лучам. Строение органа зрения играет значительную роль при их ориентировке во время движения и суточном ритме активности. Это обусловливает возможности охоты хищника за жертвой и защиты жертвы от хищника (Иванов, 1988).
Большинство рыб ведут дневной образ жизни, и поэтому свет для них имеет сигнальное значение при разыскивании пищи, бегстве от врагов, образовании стай, миграциях, созревании гонад.
По отношению к свету различают дневных (светолюбивых) и сумеречных (светобоязливых) рыб. Отношение к свету неодинаково и на различных этапах жизненного цикла рыб. Рыбец сразу после выклева долго развиваются в затемненных местах на дне, взрослые же особи держатся преимущественно вблизи берега на глубине не более 25 метров, где освещённость достаточно хорошая.
Большинство рыб, за исключением сумеречных и большинства хрящевых, обладают цветовым зрением. Цветовое зрение возможно только при высокой освещенности, когда могут функционировать колбочки.
Биологическое значение цветового зрения рыб связано с возможностью распознавания окраски водных животных и приспособления их окраски к окружающему фону. Основная функция окраски рыб защитная, однако имеет большое значение и во взаимоотношениях между полами, в стайном поведении (Касымов, 1981).
Суточные изменения интенсивности освещенности являются основной причиной суточных вертикальных миграций рыб.
Продолжительность светового дня является сигнальным раздражителем, обусловливающим нерестовую миграцию.
Освещенность оказывает влияние и на скорость созревания гонад у рыб. Солнечные лучи также влияют на обмен веществ у рыб.
Освещенность для дневных рыб наряду с чувством голода является основным фактором, обусловливающим двигательную активность (Моисеев, 1981).
2.3 Влияние гидрохимических показателей
Соленость. В воде рек, озер, морей и океанов содержится большое количество различных элементов и минеральных солей. В зависимости от количества растворенных солей различают воду пресную (до 0,5%0), солоноватую (0,5—25%о), морскую (25—40%0) и пересоленную (более 40%о).
Соленость воды влияет на рост рыб. Представители одного и того же вида обычно в морской воде растут лучше, чем в солоноватой, а в солоноватой в свою очередь более интенсивно растут некоторые пресноводные рыбы. У карповых, в том числе и рыбца, максимальная подвижность и интенсивность питания, а также наиболее быстрый рост наблюдаются при солености 3—5%о (Астанин, 1968).
С повышением солености возрастает плотность воды. Растворенные в воде минеральные соли поддерживают у рыб постоянное осмотическое давление, обеспечивающее работу всех внутренних органов: всасывание в кровь через стенки кишечника питательных веществ, а также выделение продуктов обмена. Рыбы в течение своего эволюционного развития приспособились жить в воде с более или менее постоянным солевым составом, который определяет и постоянство осмотического давления в их организме. Различия в осмотическом давлении воды разной солености являются основным препятствием перехода рыб из одной среды в другую.
Возможность обитания некоторых рыб в воде различной солености обеспечивается развитием у них осморегуляторных приспособлений, направленных на сохранение внутреннего осмотического давления (Иванов, 1988).
Большое значение в жизни рыб имеет и солевой состав воды. Соли азотной, фосфорной и кремниевой кислот (биогены) способствуют развитию первичной продукции в водоемах – прежде всего фитоплактона, а следовательно, и животных (планктонных и бентосных), служащих пищей для рыб.
Солевой состав воды оказывает на жизнь рыб и прямое влияние. Так, например, фосфор и кальций, имеющие важное значение при формировании костной ткани и синтезе белков, рыбы могут получать не только из пищи, но и непосредственно из воды. Магний, калий, натрий, серу, железо, медь, йод, фтор, молибден и другие химические элементы, необходимые для нормального роста и развития, они могут также получать из воды. Однако рыбовод должен помнить, что повышенное содержание в воде той или иной соли может оказать на рыбу вредное воздействие, а в некоторых случаях даже вызывать ее гибель. Например, определенное количество растворимых в воде закисных соединений железа совершенно необходимо для развития растений и животных, ибо железо входит в состав хлорофилла растений, крови и тканей животных. Но если в воде содержится избыточное количество закисного железа, то оно при переходе в окисную (нерастворимую) форму отнимает у воды кислород и выпадает в виде бурого осадка, который у взрослых рыб вызывает заболевание глаз, а у молоди – поражение жабр, приводящее рыбу к гибели. Подобное явление можно наблюдать и при высокой концентрации соединений азота. Так, значительное содержание в воде нитратов или нитритов смертельно для рыб (Черномашенцев, Мильштейн, 1983).
Морская вода содержит в основном хлористые, а пресная – углекислые и сернокислые соли, поэтому пресная вода бывает жесткой или мягкой.
Внесение минеральных удобрений в водоемы создает благоприятные условия для развития кормовой базы и способствует повышению их продуктивности. Фосфорные удобрения не только способствуют улучшению кормовой базы, но и непосредственно воздействуют на выращиваемую молодь рыб, повышая обмен веществ, стимулируя рост и развитие рыб.
Большое влияние на обмен веществ рыб оказывают содержащиеся в воде соли железа, которые в концентрации до 0,1 мг/л стимулируют рост рыб, а при большем содержании вызывают снижение потребления кислорода и замедление их роста.
Растворенные в воде газы. Вода как среда обитания рыб содержит растворенные газы, особенно кислород, азот и в небольшом количестве углекислый газ.
Все рыбы дышат растворенным в воде кислородом, поэтому содержание его в воде имеет для них решающее значение. Лишь немногие рыбы частично приспособились к дыханию атмосферным кислородом. К содержанию кислорода в воде рыбы относятся неодинаково. Как правило, пелагические рыбы, речные и холодолюбивые, более требовательны к содержанию кислорода, чем донные, озерные и теплолюбивые.
По отношению к содержанию кислорода в воде всех рыб можно разделить на четыре группы.
1. Рыбы, живущие в воде с высоким содержанием кислорода (7 – 8 мл/л). К этой группе рыб относятся лососевые, которые ощущают недостаток кислорода при содержании его в воде в количестве 4 – 5 мл/л.
2. Рыбы, требующие сравнительно высоких концентраций кислорода в воде (6 – 7 мл/л), но способные жить и при содержании кислорода в воде 5 – 6 мл/л. К этой группе рыб относятся осетровые.
3. Рыбы, которые могут жить при небольшом количестве кислорода в воде (4—5 мл/л). К этой группе относятся сазан, лещ, судак и др.
4. Рыбы, которые могут жить в воде с незначительным содержанием кислорода (0,5 мл/л) – золотой карась.
Рыбец является морским видом и поэтому относится ко второй группе рыб, т.е. требующих поддержание концентрации кислорода на уровне 6 – 7 мл/л. Во время нерестовых миграций эти рыбы способны жить при меньших концентрациях кислорода (Рыжов, 1987).
При недостаточном содержании кислорода в воде обмен веществ в организме рыб снижается, и это отрицательно сказывается на их росте и развитии. Поэтому для каждого вида рыб существует нижняя граница содержания кислорода в воде - так называемый кислородный порог, за пределом которого организм не в состоянии осуществлять свои жизненные функции и погибает от удушья (Иванов, 1988). Рыбец погибает при содержании кислорода в воде ниже 0,4 – 1 мл/л. Кислородный порог не всегда постоянен: он сдвигается в зависимости от температуры воды, солевого состава и концентрации водородных ионов (Привезенцев, 2004).
Количество растворенного в воде кислорода зависит от температуры, солености, ледового покрова, развития растительности, процессов распада органического вещества и др.
При повышении температуры и солености растворимость кислорода в воде уменьшается. Так, при 0° С и солености 0 %о в воде может раствориться 10,29 см3/л, а при 30° С – только 5,57 см3/л кислорода (Иванов, 1988).
С повышением температуры воды рыбы потребляют больше кислорода. Однако существует температурный порог, по достижении которого с дальнейшим повышением температуры воды потребление кислорода падает. У рыбца потребление кислорода снижается при 26 – 28 °С, а при температуре свыше 30—35 °С они могут погибнуть. При быстром повышении содержания кислорода у рыб появляются беспокойство, одышка, кислородный наркоз, и они погибают от удушья (Берлянд, 1953).
Избыток кислорода в воде по сравнению с оптимальным режимом в период эмбрионального развития снижает функцию кроветворных органов, что вызывает анемию у рыб.
Потребление кислорода рыбами зависит от их вида, возраста, подвижности, плотности посадки, физиологического состояния, также температуры и солености воды.
На потребление кислорода рыбами оказывает влияние и соленость воды. У пресноводных рыб, например, при небольшом увеличении солености обмен веществ возрастает, а при значительном замедляется, и потребление кислорода уменьшается.
Обычно молодь рыб более требовательна к содержанию кислорода, чем старшие возрастные группы.
Интенсивность обмена веществ и потребление кислорода одиночными рыбами и в скоплениях неодинаковы. При высокой плотности населения рыб потребление кислорода ими снижается. У рыб, залегающих на зимовку в ямы, потребление кислорода по сравнению с одиночными рыбами значительно уменьшается.
Потребление кислорода изменяется в зависимости от физиологического состояния рыбы. Перед нерестом у некоторых рыб потребление кислорода повышается на 25—50% первоначального.
При плохом кислородном режиме интенсивность питания низкая и не увеличивается даже при обилии корма.
Снижение содержания кислорода может привести к летним и зимним заморам (Черномашенцев, Мильштейн, 1983).
Содержание в воде свободного диоксида углерода – важный гидрохимический показатель, характеризующий пригодность водоема для рыборазведения. Углекислый газ образуется в результате дыхания животных и растений, при разложении органических веществ. Большие концентрации в воде свободного диоксида углерода могут оказаться для рыбы не только вредными, но и губительными даже при наличии достаточного количества кислорода. Даже при небольшом содержании углекислого газа в воде кровь теряет способность усваивать кислород, и рыба погибает от удушья. Если содержание углекислого газа и не вызывает гибели, то сильно замедляет рост рыбы, так как снижается усвояемость пищи. Допустимой для большинства рыб концентрацией считается 10 – 30 мл/л (Привезенцев, 2004).
Растворенный в воде азот, находящийся в свободном состоянии, не оказывает влияния на жизнь рыб и является для них индифферентным. В теле рыб содержится значительное количество азота (1,5 – 4 %), входящего в состав всех белков, синтезируемых организмом рыб из азотистых веществ потребляемой пищи, в частности из ее аминокислот.
Из других газов в воде могут присутствовать сероводород и метан, которые образуются на дне водоемов в результате гниения органических веществ без доступа воздуха. Эти газы ядовиты и губительно действуют на организм рыб. Особенно опасен для рыб сероводород, который в отличие от метана хорошо растворим в воде и может в ней задерживаться. Метан почти не растворим в воде. Он быстро поднимается со дна водоема в верхние слои воды, а затем улетучивается в атмосферу. Однако непрерывное выделение метана из грунта водоема бывает достаточным для того, чтобы при прохождении его через воду отравить рыбу, попавшую в зону действия этого газа.
Следует отметить, что иногда в некоторых участках рек с быстрым течением, особенно под водосливом гидроэлектростанций, вода перенасыщается газами воздуха, что может вызвать у рыб газопузырьковую болезнь (Рыжов, 1987).
Активная реакция среды (рН), имеющая важное значение для жизни рыб, зависит от соотношения растворенных в воде кислорода и свободной углекислоты и закономерно изменяется в зависимости от суточного и сезонного хода фотосинтеза.
Морская вода имеет щелочную реакцию (рН колеблется около 8,0). В пресных водоёмах рН изменяется от очень кислой до сильно щелочной реакции.
Это связано с тем, что в пресных водоемах избыток углекислого газа вызывает увеличение кислотности воды, в то время как в морской, содержащей в большом количестве бикарбонаты, избыток этого газа связывается, и рН более постоянна.
Для каждого вида рыб характерны определенные значения активной реакции среды. При изменении этих величин обмен веществ нарушается, так как снижается способность организма поглощать кислород. Наиболее благоприятна для жизни рыб, являющихся объектами массового искусственного разведения, нейтральная или слабощелочная реакция воды (рН 7,0—7,5). При рН ниже 6,0 и выше 8,5—9,0 рыбы могут погибнуть (Иванов, 1988).
Глава 3. Управление половыми циклами у рыбца различными методами
В практике искусственного рыборазведения применяют три метода стимулирования созревания половых продуктов у производителей рыб: экологический, физиологический и эколого-физиологический.
Экологический метод. В 30-е годы А. Н. Державин провел многочисленные опыты по выдерживанию производителей реофильных рыб в садках с речной водой и выявил те факторы, которые способствуют развитию и созреванию половых клеток, овуляции и образованию спермы. Это прежде всего течение, кислородный режим и галечный грунт (нерестовый субстрат). Он установил также, что при этом очень важно поддерживать температуру, близкую к температуре нереста данного вида рыб.
В настоящее время экологический метод широко применяется в практике искусственного разведения лососевых и реофильных карповых рыб при выдерживании производителей с целью получения от них зрелых половых продуктов. Также экологический метод применяется при искусственном разведении осенне – нерестующих рыб, входящих в реки с гонадами во ІІ, ІІ – ІІІ, ІІІ стадиях зрелости (Привезенцев, 2004).
Именно этот метод используется при искусственном разведении рыбца. Производителей рыбца заготовляют для искусственного разведения весной в период нерестового хода в реки (март – апрель), где их ловят неводами. Отловленных производителей осматривают и отбирают более крупных особей, не имеющих травм, с учетом соотношения самок и самцов 2:1. Отобранных производителей сажают в прорезь. Прорезь транспортируют буксиром на рыбоводный завод. При этом скорость транспортировки не должна превышать 3—4 км/ч (Рыжов, 1987).
На рыбоводном заводе производителей выгружают из прорези, помещают в заполненные водой брезентовые чаны, или носилки, и доставляют к садкам, в которые пересаживают их на выдерживание до созревания половых продуктов.
Требуемое заводу количество производителей заготавливают с учетом возможного отхода 10 % особей в период выдерживания в садках и несозревания 20 % особей.
Садки для выдерживания производителей расположены в наименее шумном месте завода. Они стационарные и земляные. В верхней части каждого садка (пруда) имеется по три нерестовых канавы, а в нижней части – водоспуск (рис. 10). Ширина садка – 12 м, длина – 35 м (без канав), глубина –0,5 – 1 м. Нерестовые канавы делают длиной 25 м и шириной по дну 0,8 м. Дно и откосы их покрыты гравием и ракушкой слоем 15 – 20 см. Дно канав имеет уклон в сторону садка. Глубина в верхней части канав – 15 см, а в нижней –45 см. Канава имеет четыре переката, на которых уложена галька слоем в 3 –7 см. Через каждые 5 м канава разделена съемными решетками на отсеки. Вода поступает из отстойника в канал, из которого она подается в канавы. Это обеспечивает подачу не мутной воды и отсутствие резких температурных ее колебаний, что наряду со скоростью ее течения в канавах не менее 0,5 – 0,7 м/с положительно влияет на созревание половых продуктов производителей рыбца.
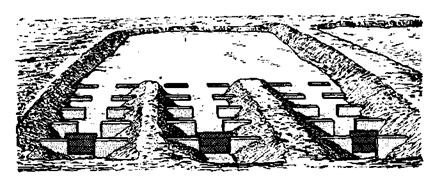
Рис. 10. Садок для выдерживания производителей рыбца
Расход воды в садке составляет 60 – 85 л/с, а плотность содержания в нем производителей может быть до 5 особей на 1 м2.
При наступлении нерестовых температур производители рыбца по мере созревания гонад выходят из садков в нерестовые канавы. Массовый заход производителей в эти канавы происходит при температуре воды 18 °С. За зашедшими в канавы производителями ведут наблюдение. При обнаружении готовности производителей к нересту секции канав перегораживают решетками, уменьшают подачу воды и отлавливают их. От зрелых особей берут икру и сперму, а затем совместно с еще недозревшими рыбами их вновь сажают в садок для получения второй порции половых продуктов (Берлянд, 1953).
Кроме указанного метода получения зрелых производителей рыбца, применяют метод выдерживания этой рыбы совместно с шемаей (Астанин, 1968).
Физиологический метод. В 30-е годы Н. Л. Гербильский осуществил многочисленные гистологические исследования гипофиза рыб и раскрыл механизм его физиологического воздействия на созревание половых продуктов.
В естественных условиях переход рыбы в нерестовое состояние осуществляется при наличии совокупности определенных факторов внешней среды. Они воспринимаются органами чувств рыбы, а через них действуют на ее центральную нервную систему – на гипоталамус. Клетки гипоталамуса выделяют гормон, активизирующий гормональную деятельность гипофиза. Выделяемый гипофизом гонадотропный гормон поступает в кровь и стимулирует созревание половых клеток, а также выход зрелых яиц (икринок) из фолликул и образование спермы. Во время нереста производители рыб выметывают зрелые половые продукты не все сразу, а постепенно. Так, самка выметывает икринки в воду по мере их овуляции. Совершаемые самкой движения приводят к разрыву следующих фолликул и продолжению вымета икринок.
Н. Л. Гербильский установил гонадотропную активность гипофиза рыб в различные периоды годового цикла. Гонадотропный гормон поступает в кровеносную систему организма рыбы непостоянно и в различном количестве. В определенные сезоны года он накапливается в гипофизе. Это позволяет использовать гипофиз как источник гонадотропного гормона, при помощи которого можно получать зрелые половые продукты от производителей на рыбоводных предприятиях. При внутримышечных инъекциях производителям суспензии гипофиза рыб гонадотропный гормон поступает в кровь и стимулирует половой процесс. Это приводит к быстрому переходу половых желез производителей из IV в V стадию зрелости и получению от них зрелой, способной к оплодотворению и развитию икры у самок и доброкачественной спермы у самцов. Следовательно, при искусственном рыборазведении можно получать зрелые половые продукты от производителей путем инъецирования им препарата гипофиза. В этом случае созревание половых клеток, овуляция и образование спермы происходят, как и при естественном нересте, под влиянием гипофиза. Однако отличие состоит в том, что увеличение количества гонадотропного гормона гипофиза в крови производителей происходит не под влиянием нерестовых условий, усиливающих выделение собственного гонадотропного гормона, а при помощи введения им взятых гипофизов от других рыб (Привезенцев, 2004).
Н. Л. Гербильский и Б. Н. Казанский установили, что для стимулирования созревания половых продуктов у разных объектов рыбоводства можно пользоваться гипофизами различных рыб. Однако в ряде случаев гонадотропный гормон обладает видовой специфичностью. Так, гонадотропный гормон судака и окуня не вызывает созревания половых продуктов у карповых. В связи с этим при проведении гипофизарных инъекций нужно использовать гипофизы тех видов, которые содержат эффективный гонадотропный гормон. Для стимуляции созревания половых продуктов у производителей можно применять гипофизы, взятые у того же вида рыб. Гонадотропный гормон сазана стимулирует созревание половых продуктов у производителей многих видов рыб, относящихся к различным семействам. Но на рыбоводных предприятиях гипофизы сазана обычно вводят производителям семейства карповых – сазану, карпу, белому амуру, белому и пестрому толстолобикам. Гипофизарные инъекции прочно вошли в практику рыбоводства и особенно широко применяются на осетровых рыбоводных заводах.
Для многих карповых, в том числе и рыбца, физиологический метод практически не применим. Это связано с тем, что количество производителей, от которых получают половые продукты, достаточно велико и провести инъецирование каждого затруднительно по техническим причинам. К тому же это требует значительных финансовых затрат (Иванов, 1988).
Эколого-физиологический метод. Советские ученые разработали и внедрили в производство эколого-физиологический метод, который предусматривает стимулирование созревания половых продуктов у производителей путем комбинированного воздействия на организм рыбы экологических факторов среды и вводимых физиологически активных веществ.
В настоящее время этот метод широко используется на наших рыбоводных предприятиях для усиления стимулирующего воздействия на производителей с целью ускорения созревания половых продуктов и повышения эффективности применения гипофизарных инъекций (Астанин, 1968).
Следовательно, на современном этапе развития искусственного рыборазведения эколого-физиологический метод сочетает гипофизарные инъекции с выдерживанием производителей в условиях, близких к естественным. Это дает возможность рыбоводу получать в определенный день и даже час необходимое количество зрелой икры и спермы, что позволяет планировать работу рыбоводного предприятия по каждому звену биотехнического процесса.
Глава 4. Биологические основы кормления рыбца
Все вещества, необходимые для нормального развития, рыба получает с пищей. Потребление пищи начинается сразу после рассасывания у личинок желточного мешка на две трети. В этот период рыбы переходят на так называемое смешанное питание. После израсходования запасов желточного мешка рыба переходит на внешнее, т.е. активное, питание ( Иванов, 1988)
В естественных условиях пищевые потребности рыб удовлетворяются за счёт животных и растительных организмов, имеющихся в водоёме.
При искусственном выращивании рыб содержат в условиях уплотнённых посадок, и пищевых ресурсов водоёма не хватает. Поэтому рыбу кормят вносимыми кормами, подразделяемые на живые и не живые или искусственные (Привезенцев, 2004).
Рыбца чаще всего выращивают совместно с другими карповыми рыбами (в частности сазаном, карпом, белым амуром), как добавочный объект и поэтому при кормлении чаще всего применяют те же корма, что и для карпа (Рыжов, 1987). Для интенсификации рыбоводного процесса целесообразно использовать естественную кормовую базу и живые корма, так как они являются наиболее полноценными, содержат все необходимые питательные вещества и охотно поедаются рыбами. В настоящее время на рыбоводных предприятиях широко используют искусственное разведение коловраток, ракообразных (дафнии, моины, артемии, гамариды и т.д.), олигохет, личинок насекомых.
Разводимые кормовые организмы обладают высокой пищевой ценностью, быстрым созреванием и ростом, высокой плодовитостью. При выращивании кормов и их использовании нужно широко применять механизацию трудоёмких процессов, что позволяет добиться значительного экономического эффекта (Карпевич, 1981).
В естественных условиях спектр питания рыбца значительно изменяется в течении его развития от личинки до взрослой рыбы. Питание начинается на втором этапе личиночного периода развития. В это время личинки питаются одноклеточными водорослями, коловратками, инфузориями, молодью кладоцер и копепод. На пятом этапе развития личинки переходят на питание взрослыми формами кладоцер и копепод,личинками фитофильных хирономид. К концу малькового периода развития рыбцы питаются, как и взрослые рыбы, зоопланктоном и зообентосом: мелкими моллюсками, личинками насекомых, червями, бокоплавами (Астанин, 1968).
Технологии кормления молоди карповых рыб созданы учёными ВНИИПРХ (д.б.н. Гамыгин Е.А., к.б.н. Боева Т.М.) и НТЦ «Астаквакорм» (д.б.н Пономарёв С.В.), технологии кормления товарной рыбы – специалистами КрасНИИРХ, ГосНИОРХ, ВНИИПРХ, УкрНИРХ.
При уплотненных посадках для кормления рыбца необходимо использовать кормовые смеси, которые должны удовлетворять ряду условий. Так, в составе смесей из нескольких компонентов количество каждого не должно превышать 60 %• от всего корма при содержании не менее 10 – 15% связующих веществ. Корм должен быть физиологически полноценным, т. е. сбалансированным по аминокислотному составу, витаминам, минеральным веществам. Поскольку большинство компонентов, особенно жмыхов, не содержит полного набора аминокислот и нуждается в обогащении, в состав смеси должно входить не менее двух видов жмыхов. При выращивании рыбца его кормят различными кормовыми смесями (комбикормами), изготовленными с учетом физиологических особенностей рыб различных возрастов, сбалансированными по всему комплексу питательных веществ и обогащенными рядом добавок (Скляров и др., 1984).
Все рыбы, в том числе и рыбец, отличаются высокой потребностью в белке. Общими для всех белков являются 24 аминокислоты, однако полноценность белка определяется наличием и соотношением в нём незаменимых аминокислот, а также их доступность для организма рыбы. Незаменимыми для рыб являются 10 аминокислот: лизин, метеонин, трептофан, аргинин, гистидин, фенилаланин, треонин, валин, лейцин, изолейцин. Сбалансированный по аминокислотному составу корм имеет решающее значение для роста рыбы, а также для сведения к минимуму загрязнения воды азотом, т.к. при окислении аминокислот азот выделяется в виде аммиака. Потребность рыбца в протеине зависит от возраста и условий выращивания. При кормлении рыбца содержание протеина в стартовых комбикормах должно быть не менее 45%. При выращивании молоди, товарной рыбы, а также производителей количество белка должно быть 23 – 32% (Астанин, 1968).
Изменение температурного режима также влияет на потребность рыбца в белке. С повышением температуры воды увеличивается скорость протекания обменных процессов и тем самым степень усвоения белка (Берлянд, 1953).
Основными источниками белка в комбикормах являются: рыбная мука, мясная мука, мясокостная мука, крилевая мука, мука из куколок тутового шелкопряда, рыбный фарш, молочные продукты, дрожжи и др.
Важную роль в энергетическом обмене играют жиры. Липиды являются источником незаменимых жирных кислот, с ними связано поступление и накопление в организме жирорастворимых витаминов, таких как ретинол, альфа-токоферол. Наиболее эффективно вводить в корма жиры содержащие ненасыщенные жирные кислоты: пальмитолеиновая, леиновая, ленолевая, леноленовая, архидоновая. Содержание жиров в комбикормах для рыбца не должно превышать 12 %. Основными источниками жиров являются: рыбий жир, рыбная мука, растительные масла.
Углеводы – простейшие из основных энергосодержащих компонентов кормов. Оптимальный уровень содержания углеводов в кормах для рыбца 40 – 50 %; в индустриальных условиях – для сеголеток массой 1 – 150 г 29 – 37 %, для товарной рыбы 33 – 44% (Зайцев, 1988).
В комбикорма для рыбца необходимо добавлять и такие вещества, которые, не являясь структурными элементами и не поступая в качестве источников энергии, оказывают большое влияние на обмен веществ. Это витамины, минеральные вещества, которые вводятся в корма в виде витаминно-минеральных премиксов. Также в корма добавляют пищевые красители, атрактанты, антибиотики, наполнители и другие компоненты.
Все смеси готовят в виде теста, гранул или брикетов. При гранулировании кормов их меньше расходуется на единицу прироста рыбы, они меньше теряют в воде питательных веществ, содержат все необходимые добавки и меньше загрязняют водоем органическими веществами. Гранулирование позволяет использовать механизацию при приготовлении кормовых смесей, транспортировке и внесении кормов в пруды.
Эффективность кормления значительно возрастает при использовании брикетированных кормов, которых расходуется при выращивании рыбы на 12 — 15 % меньше, чем тестообразных.
Начало кормления рыбца приурочивают к повышению температуры воды до 12 °С. Для кормления личинок используют стартовый комбикорм РК-С3М. Основу этого корма составляют высокобелковые продукты микробиосинтеза, обезжиренная рыбная мука, казеинат натрия, растительное масло, пщеничная мука и поливитаминный премикс (Скляров и др., 1984).
Суточная норма кормления определяется температурой воды и массой личинок (табл.1)
В аппаратах ВНИИПРХ периодичность кормления составляет 0,5 часов, в других рыбоводных ёмкостях – не реже 1 часа. При использовании автоматических кормораздатчиков периодичность кормления составляет до 0,2 – 0,3 часов. Кормление молоди проводят в течении светового дня. Разовую порцию корма разбрасывают равномерно по поверхности воды в местах скопления личинок (Зайцев, 1988).
Корм РК-С3М рассчитан на использование в условиях недостатка естественной пищи. Однако по возможности следует способствовать попаданию мелких форм зоопланктона в рыбоводные ёмкости. Наличие даже минимального количества живых кормовых организмов в пище молоди способствует скорости роста и общему улучшению рыбоводных показателей.
Таблица 1
Суточная норма кормления личинок и мальков карповых рыб, % от массы тела
| Масса личинок, мг |
Температура воды, °С |
||
| 20 – 25 | 25 – 28 | 29 – 32 | |
| До 3 | 50 | 50 | 50 |
| 3 – 10 | 50 | 60 | 75 |
| 10 – 50 | 70 | 90 | 80 |
| 50 – 100 | 50 | 70 | 80 |
| 100 – 300 | 40 | 50 | 60 |
| 300 – 1000 | 25 | 30 | 40 |
| 1000 – 2000 | 15 | 20 | 30 |
Для кормления сеголетков рыбца массой от 1 до 50 г используется комбикорм АК-1КЭ, который состоит рыбной муки, мясокостной муки, жрожжей, соевого шрота, масла растительного, премикса, дикальция фосфата (Скляров и др., 1984).
Для кормления рыбца от 50 г до товарной массы используют экструдированный комбикорм АК-2КЭ. Размер гранул должен соответствовать массе выращиваемой рыбы (табл.2).
Таблица 2
Рекомендуемые соотношения между размером гранул (крупки) и массой рыбца
| Масса рыбы, г | Размер гранул, мм |
| 1 –10 | 1,5 – 2,5 |
| 10 – 40 | 2,5 – 3,5 |
| 40 – 150 | 3,5 – 4,5 |
| 150 – 500 | 5 – 6 |
| Более 500 | 6 – 8 |
Зимой при температуре воды свыше 6 оС кормление рыбы не следует прекращать, при этом суточный рацион должен быть не высоким и обеспечивать лишь поддерживающий обмен. При температуре воды 6 – 8 оС суточная норма составляет до 0,5 % , при температуре 9 –10 оС – до 1% , при температуре 11 – 12 – до 2% от массы тела. В зимний период года, при низком уровне обмена веществ, следует использовать низкобелковые кормосмеси.
После привыкания рыбы к корму величину рациона рассчитывают в зависимости от прироста и уточняют через каждые 10 суток. В период выращивания рыбы рацион уточняют по результатам контрольных обловов (Скляров и др., 1984).
Эффективность использования кормов во многом зависит от условий их хранения. Рассыпные корма хранят в сухих помещениях—складах, в которых пол изолирован от почвы водонепроницаемыми материалами. Комбикорм ссыпают в бурты высотой до 1,5 м и шириной 10—15м и хранят не более 2мес. При перевозке рассыпных и гранулированных кормов кузова автомашин или тракторных прицепов выстилают изнутри и накрывают сверху брезентом. Чтобы устранить потери, неизбежные при дополнительных перевалках, рассыпные и гранулированные корма рекомендуется хранить в башнях силосного типа на прудах на центральных усадьбах. При использовании таких хранилищ на прудах загрузка кормораздаточных средств производится самотеком.
Рассыпные корма вносят в пруды в тестообразном состоянии, для чего на небольших прудах используют лодки, а на больших водоёмах – кормораздатчики разных конструкций.
Кормление рыб проводят с учетом их возрастных особенностей и термического и гидрохимического режимов водоемов. Молодь начинают кормить с двух-трехнедельного возраста. Кормление двухлетков начинают в период, когда температура воды повышается до 8 –12 °С. Производителям и ремонтному стаду корм задают из расчета 2,5 – 3,0 % от массы тела с учетом поедаемости. Корм вносят на кормовые точки или кормовые дорожки, отмеченные вешками.
Гранулированные корма лучше вносить на специальные кормушки, которые бывают подъемными, самовсплывающими и автоматическими.
Подъемные кормушки делают в виде квадратных деревянных площадок размером 0,8X0,8 м, окруженных бортиками высотой 15 см. Через отверстие в центре площадки пропущен шест, забиваемый в дно пруда. Площадки закрепляют на шесте клиньями.
Самовсплывающая кормушка состоит из металлического столика с дном и верхним и нижним обручами, четырех стоек из проволоки, приваренных к обручам. К верхнему обручу крепят пенопластовые поплавки, а к нижнему – груз массой 2 – 3 кг. Для удержания столика на одном месте поплавок подвязывают к верхнему обручу бечевкой, имеющей длину, равную глубине водоема.
Автоматические кормушки представлены устройством НЕ-2 (производство ВНР) и кормушками «Рефлекс» разных моделей, различающихся вместимостью бункера. Устройство НЕ-2 представляет собой бункер, укрепленный с помощью телескопических стоек к поддону шириной до 2 м. Корм высыпается на поддон по мере его поедания. В один бункер можно засыпать до 6 ц сухих кормов, преимущественно гранулированных. Одна кормушка обслуживает 2,9 га прудов.
Раздачу корма проводят в одни и те же часы. При одноразовом кормлении корм задают в 7 – 9 ч, при двухразовом – в 7 – 9 и 13 – 14 ч, а при трехразовом – в 6 – 8, 11 – 13 и 16 – 17 ч. Через 2 ч после внесения корма в пруд с помощью специального черпака проверяют его поедаемость (Зайцев, 1988).
Глава 5. Транспортировка икры, личинок, молоди и взрослых особей рыбца
При работах по воспроизводству рыбных запасов широко применяют перевозки икры, личинок, молоди и производителей. Транспортировка половых продуктов, посадочного материала и взрослой рыбы может быть внутрихозяйственной и межхозяйственной. Непродолжительные по времени внутрихозяйственные перевозки осуществляют в полиэтиленовых метках, живорыбных контейнерах, молочных флягах и живорыбных автомашинах (в зависимости от вида и возраста рыб, а также расстояния перевозки). Для межхозяйственных перевозок на большие расстояния используют авиацию, железнодорожный и автомобильный транспорт (Иванов, 1988).
Перевозка спермы. В связи с тем, что в семенной жидкости сперматозоиды находятся в активном состоянии, сперму рыбца можно перевозить на любые расстояния в сухих стерильных пробирках, установленных в термосе со льдом. При этом необходимо учитывать сроки ее активности. Длительность оплодотворяющей способности спермы рыбца при температуре 0 – 2 оС составляет 6 – 8 суток, при 2 – 5 оС – 2 суток. Хранению и перевозке подлежит свежеотобранная сперма, помещенная в сухие пробирки отдельно для каждого самца с плотно пригнанными пробками во избежание попадания воды.
Пробирки с этикетками заворачивают в марлю и отпускают в термос, наполненный мелко наколотым льдом.
Перевозка икры. Для перевозки неоплодотворенной икры ее закладывают в сухую банку, которую плотно закрывают пробкой и помещают в термос. Банка должна полностью заполняться икрой без свободного воздушного пространства.
Перевозка оплодотворенной икры каждого вида рыб осуществляется на определенной стадии развития эмбриона. Для рыбца наиболее желательна перевозка на стадии пигментации глаз.
При кратковременной транспортировке обесклеенную икру перевозят в банках, в которые наливают воду, а затем постепенно заполняют икрой. Наполнив банки доверху икрой, сцеживают воду через марлю. Затем банки с икрой ставят в изотермический ящик, в котором сохраняется необходимая температура при транспортировке. Для предохранения икры от тряски банки перекладывают бумагой, ватой, поролоном или другим мягким материалом.
При длительной транспортировке обесклеенной икры используют различную тару для перевозки икры. Икру рыбца перевозят на деревянных рамках, уложенных в изотермические, влагонепроницаемые пенопластовые ящики. Рамки размером 34x28 см разделены пополам планками и обтянуты сеткой из синтетического материала. На каждой рамке положена влажная марлевая салфетка размером 70x50 см, на которой в 1,5—2 слоя разложена икра.
Икру раскладывают на рамки в воде. Для этой цели применяют заполненный водой лоток. На рамки расстилают марлевые салфетки. Уложенную на рамку икру накрывают свободными концами салфетки. Рамки в количестве 20 шт., на которых размещено около 250 тыс. икринок, укладывают в стойку ящика. При температуре воздуха выше 7 оС на дно ящика предварительно ставят пенопластовую кювету с высотой бортиков 5 см. Сверху на стопку рамок с икрой ставят такую же кювету, но с сетчатым дном. В эту кювету закладывают лед. Ящик с икрой закрывают крышкой и перевозят к месту назначения. За время транспортировки икры лед постепенно тает. Такая вода стекает по стопке рамок, охлаждая и увлажняя икру, и попадает в нижнюю кювету, которая является водоприемником (Понамарева, Бахарева, 2002).
Весьма удобной тарой для перевозки икры рыбца является картонный или фанерный ящик с уложенными в него кюветами из пористого стиролового пластика. Высота бортиков с внешней стороны кюветы равна 6,8 см, а с внутренней стороны 6 см. Толщина бортиков по верху 1,5 см. С внешней стороны бортики прямые, а с внутренней стороны с уклоном ко дну. В связи с этим внутренняя рабочая часть кюветы имеет размеры 26 × 26 см по низу и 30 × 30 см по верху. В дне кюветы 7 рядов круглых отверстий, диаметр которых не превышает размер икринок. В кювету закладывают икру, размещая её в 7 слоев на влажной марлевой салфетке. Концами салфетки прикрывают икру. Ящик вмещает 7 уложенных в стопку кювет, из которых 5 с икрой, 1 со льдом (верхняя) и одна без отверстий (нижняя), предназначенная для приема стекающей воды из кюветы с икрой, лежащей над ней. Эта вода образуется при таянии льда на верхней кювете и проходит по всем 5 кюветам, увлажняя и охлаждая на своем пути икру. Материал, из которого изготовлены кюветы, отличается легкостью и высокими изоляционными свойствами. Это позволяет поддерживать температуру при транспортировке икры не выше 7°С (Рыжов, 1987).
Если икру перевозят поздней осенью или зимой при низких температурах на транспорте, то в тару не только не закладывают лед, но, наоборот, ее утепляют, чтобы избежать промерзания икры. Стопку рамок (кюветов) обертывают плотной бумагой и перевязывают шпагатом. Для доступа воздуха бумагу, закрывающую рамки, в нескольких местах прорывают. Упакованную в бумагу стопку рамок опускают в пенопластовый изотермический ящик (Черномашенцев, Мильштейн, 1983).
В каждый транспортировочный ящик вкладывают конверт с накладной, в которой указаны количество икры, дата оплодотворения и температура инкубации.
На крышке транспортировочного ящика делают надпись «Живая икра. Верх. Не кантовать». С боковой стороны ящика прикрепляют этикетку с адресом назначения и отправления (Кожин, 1971).
Необесклеенную икру перевозят без воды во влажной среде. При кратковременных перевозках приклеенную к субстрату икру помещают в картонную коробку, дно которой устлано полиэтиленовой пленкой или мхом, прикрытым мокрой марлевой салфеткой. При отсутствии мха дно коробки можно застелить стеблями жесткой надводной растительности: рогоза, тростника, камыша. В пути субстрат с икрой периодически опрыскивают водой. В жаркие дни на дно коробки кладут небольшие кусочки льда, которые предохраняют икру от нагревания и быстрого обсыхания. При продолжительной транспортировке икру перевозят в таре, предназначенной для длительной перевозки обесклеенной икры. Субстрат с приклеенной икрой размещают на рамках и прикрывают влажными марлевыми салфетками. 6 – 8 рамок кладут стопкой в транспортировочный ящик из пенопласта. Для поддержания невысокой температуры (ниже 8°С) сверху стопки рамок устанавливают пенопластовую кювету со льдом, а снизу ставят пустую кювету (Понамарева, Бахарева, 2002).
При перевозке обесклеенной и необесклеенной икры во влажной среде при температуре 4 – 7°С весной и осенью и 8 – 12 °С летом ее отход за 24 – 48 ч транспортировки обычно не превышает 2 %.
Весьма удобной тарой для перевозки предличинок, личинок и молоди рыбца являются полиэтиленовые пакеты (рис. 11). Вместимость стандартного полиэтиленового пакета – 40 л. Их заполняют на 0,5 объема водой и предличинками (или личинками, или молодью), а оставшееся свободное пространство заполняют чистым кислородом, который подается под давлением из баллона. Закрывают зажимом Мора и устанавливают в ящики из картона (Понамарева, Бахарева, 2002). Содержащийся в пакетах кислород постепенно проникает в воду и насыщает ее. Это дает возможность транспортировать предличинки, личинки и молодь рыб при плотных посадках. Однако для принятия решения о перевозке предличинок рыбца в этих пакетах необходимо знать их биологические особенности и поведение на той или иной стадии развития. Это во многом предопределяет успех перевозки. При транспортировке необходимо поддерживать благоприятную температуру воды, для карповых в том числе и рыбца не выше 25 °С. Если рыб перевозят в жаркие дни, то в транспортировочные ящики закладывают такую массу льда, которая обеспечивала бы поддержание благоприятной температуры для рыбы (Иванов, 1988).
При расчете плотности посадки предличинок, личинок и молоди рыбца в герметически закрытый стандартный полиэтиленовый пакет, заполняемый водой и кислородом, необходимо учитывать следующее:
- степень растворения кислорода в воде в зависимости от механического перемешивания в пути, температуры и парциального давления;
- пороговое содержание кислорода для предличинок, личинок и молоди (1 – 3 мл/л);
- потребление кислорода и выделение свободной углекислоты предличинками, личинками и молодью в зависимости от температуры и их массы;
- предельно допустимые концентрации углекислоты для предличинок, личинок и молоди;
- коэффициент растворения углекислоты в воде в зависимости от температуры;
- фактор пространственного расположения предличинок, личинок и молоди (для рыбы до 1 г принимается соотношение ее массы и воды от 1 : 8 до 1 : 10, а свыше 1 г – от 1 :2 до 1:6);
- длительность перевозки предличинок, личинок и молоди.
Нормативы плотности посадки предличинок, личинок и молоди карповых, в том числе и рыбца в полиэтиленовые стандартные пакеты представлены в табл.3.

Рис. 11. Полиэтиленовый пакет для перевозки живой рыбы
Перевозка предличинок, личинок и молоди рыбца в полиэтиленовых пакетах удобнее и экономичнее, чем в других емкостях. В этих пакетах предличинки, личинки и молодь рыбца можно отправлять прямыми рейсами самолетов без сопровождающих. Пакеты, доставленные к месту назначения, помещают на некоторое время в водоем или заполненный водой бассейн, в который будет выпущена рыба. После того как температура воды в пакетах и в водоеме (бассейне) становится одинаковой, пакеты вскрывают и выпускают привезенный материал. При соблюдении указанных норм загрузки пакетов предличинками, личинками и молодью рыб отход их за период транспортировки обычно не происходит.
Таблица 3
Плотность посадки предличинок, личинок и молоди рыбца в стандартный пакет, шт.
| Температура воды, °С | Масса особи, г | Длительность транспортировки, ч | ||
| 10 | 25 | 50 | ||
| 1 | 2 | 3 | 4 | 5 |
| 5 | 5,0 | 760 | 760 | 440 |
| 5 | 10,0 | 500 | 410 | 240 |
| 5 | 20.0 | 300 | 280 | 170 |
| 10 | 1,0 | 2000 | 1900 | 900 |
| 10 | 2,0 | 1500 | 950 | 450 |
| 10 | 5,0 | 760 | 500 | 280 |
| 10 | 10,0 | 500 | 250 | 140 |
| 10 | 20,0 | 300 | 175 | 95 |
| 15 | 0,2 | 3000 | 3000 | 3000 |
| 15 | 0,5 | 2600 | 2200 | 1240 |
| 15 | 1,0 | 2000 | 1500 | 800 |
| 15 | 2,0 | 1500 | 750 | 400 |
| 15 | 5,0 | 760 | 420 | 220 |
| 15 | 10,0 | 460 | 210 | 110 |
| 15 | 20,0 | 255 | 120 | 60 |
| 20 | 0,0015 | 55000 | 55000 | - |
| 20 | 0,02 | 25000 | 25000 | 15500 |
| 20 | 0,03 | 17000 | 17000 | 10300 |
| 20 | 0,2 | 3000 | 3000 | 2300 |
| 20 | 0,5 | 2600 | 1840 | 920 |
| 20 | 1,0 | 2000 | 1000 | 550 |
| 20 | 2,0 | 1250 | 500 | 275 |
| 20 | 5,0 | 680 | 320 | 165 |
| 20 | 10,0 | 340 | 160 | 85 |
| 20 | 20,0 | 220 | 100 | 55 |
| 25 | 0,0015 | 55000 | 55000 | - |
| 25 | 0,02 | 25000 | 25000 | 15000 |
| 25 | 0,03 | 17000 | 17000 | 10000 |
| 25 | 0,2 | 3000 | 3000 | 2000 |
| 25 | 0,5 | 2600 | 1600 | 800 |
| 25 | 1,0 | 2000 | 1000 | 500 |
| 25 | 2,0 | 1150 | 500 | 250 |
| 25 | 5,0 | 760 | 300 | 160 |
| 25 | 10,0 | 400 | 150 | 80 |
| 25 | 20,0 | 205 | 95 | 50 |
Перевозка производителей рыб. Для транспортировки нескольких производителей на длительные расстояния можно использовать крупные полиэтиленовые пакеты, заполненные водой и кислородом. Для массовой перевозки производителей применяют автотранспорт, живорыбные вагоны, а также прорези. На автомашинах перевозят крупный посадочный материал на расстояния от 10 до 1000 км. Для перевозки рыбы применяются живорыбные цистерны и установки различных модификаций (АЦЖР - 3, АЦПТ - 2,8/ЗЗА, ИКА, ИКА - 4, ИКФ - 4, ИКФ - 5 и другие), смонтированные на базе автомобилей ЗИЛ - 164, ЗИЛ - 130. Они имеют емкости объемом до 3000 л, в которых поддерживаются необходимые температурный и газовый режимы.
При перевозке живой рыбы на короткие расстояния (до 50 км) отношение ее массы к массе воды должно находиться в пределах 1 : 2, при более длительной перевозке соответственно 1 : 4. (Понамарева, Бахарева, 2002). Норма загрузки карповых рыб, в том числе и рыбца устанавливается в зависимости от массы и длительности транспортировки (табл. 4).
Живорыбные вагоны используют для транспортировки рыбы на расстояния более 1000 км.
Для поддержания в транспортировочных емкостях удовлетворительных условий содержания рыбы необходима постоянная аэрация воды. При длительных перевозках транспорт помимо основного компрессора должен иметь запасной автономный бензокомпрессор или запас баллонного кислорода. Оптимальным содержанием кислорода в воде во время перевозки рыбы считается 7 – 8 мг/л, его снижение до 3 мг/л свидетельствует о критическом состоянии рыбы.
В большинстве специализированных транспортных средств предусмотрена термоизоляция. Перевозка рыбы в живорыбных вагонах может осуществляться при температурах воздуха от -40 до +30 °С.
Таблица 4
Плотность посадки карповых рыб(кг) в живорыбную машину
| Средняя масса особи,г | Темпенатура воды, °С | |||||||||
| 10 | 15 | 20 | 10 | 15 | 20 | 5 | 10 | 15 | 20 | |
| в течение,ч | ||||||||||
| 20 | 20 | 25 | 20 | 25 | 30 | 25 | 25 | 30 | 40 | |
|
При 6мг О2 / л |
При 7мг О2 / л |
При 8мг О2 / л |
||||||||
| 0,5 | - | 100 | 50 | 170 | 80 | 40 | 170 | 160 | 40 | 15 |
| 1 | - | 110 | 60 | 180 | 90 | 40 | 180 | 110 | 40 | 15 |
| 5 | - | 140 | 70 | 230 | 110 | 50 | 230 | 130 | 60 | 25 |
| 10 | - | 150 | 80 | 260 | 120 | 50 | 260 | 150 | 60 | 25 |
| 20 | 310 | 160 | 80 | 270 | 130 | 60 | 310 | 170 | 60 | 25 |
| 40 | 350 | 180 | 90 | 290 | 140 | 60 | 370 | 180 | 70 | 35 |
| 80 | 380 | 200 | 100 | 320 | 150 | 70 | 440 | 200 | 80 | 35 |
| 100 | 400 | 210 | 110 | 340 | 160 | 70 | 460 | 210 | 85 | 40 |
| 200 | 430 | 210 | 110 | 360 | 170 | 80 | 470 | 320 | 85 | 45 |
| 500 | 480 | 250 | 130 | 400 | 190 | 90 | 540 | 270 | 100 | 50 |
| 1000 | 530 | 280 | 140 | 450 | 220 | 100 | 630 | 290 | 110 | 55 |
Дальние перевозки рыбы и икры осуществляют также с помощью самолетов (Привезенцев, 2004).
Часто для перевозки производителей рыбца используют специальные суда - прорези. Это вид транспортировки самый простой и экономичный. В то же время она связана с возможностью заражения при контактировании с забортной водой. Норма посадки производителей рыбца в прорези с полезным объёмом 30 м3 составляет 7000-7700 штук.
Глава 6. Биологические основы акклиматизации рыбца
Акклиматизация рыб и кормовых беспозвоночных является составной частью комплексных мероприятий по воспроизводству рыбных запасов и кормовых ресурсов в водоемах.
Задачей акклиматизационных работ являются повышение продуктивности и хозяйственной ценности водоемов, улучшение видового состава фауны, а также сохранение и увеличение численности отдельных ценных видов водных организмов за счет расширения ареала.
Теоретические основы акклиматизации водных организмов разработаны советскими и зарубежными учеными, среди которых ведущими являются Л. А. Зенкевич, Б. С. Ильин, Б. Г. Иоганзен, А. Ф. Карпевич, Т. С. Расе (Привезенцев, 2004).
В настоящее время рыбец водятся во многих водохранилищах, созданных на реках, в последние десятилетия на территории Польши – в бассейне Вислы, в Чехословакии – в бассейне Дуная, а также в водохранилищах на реках Няманус, Даугава, Днепр, Днестр, Дон и другие. Однако большое хозяйственное значение рыбец может иметь в водохранилищах, которые как указывает П. А. Моисеев, (1981):
- достаточно большие по площади;
- имеют притоки с каменистыми и гравилистыми грунтами и быстрым течением;
- в которых не производится интенсивный лов мелкоячеистыми орудиями лова;
К таким следует отнести водохранилища в бассейне рек Дон, Няманус, Висла и другие.
Для акклиматизации объекта было выбрано Сенгилеевское водохранилище, так как этот водоём отвечает всем требованиям необходимых для эффективной полноцикловой акклиматизации: достаточная приёмная ёмкость водоёма, наличие мест для нереста рыбца, большой размер водохранилища, присутствие в значительных количествах естественной кормовой базы, благоприятный гидрохимический режим водоёма, возможность дальнейшего промыслового использования популяции рыбца. К тому же опыт предыдущих работ по акклиматизации рыбца в этом водоёме говорит о перспективности данного направления.
С точки зрения биологии вида целесообразность акклиматизационных мероприятий подтверждается тем, что рыбец является проходной и полупроходной рыбой, имеющей широкий ареал обитания, а, следовательно, приспособлен к значительным колебаниям температуры воды, солености, рН и других показателей. Рыбец хорошо приспосабливается к новым условиям среды и обладает достаточно высокой жизнестойкостью и конкурентоспособностью по отношению к другим обитателям водоёма. Необходимость мероприятий обусловлена тем, что численность рыбца повсеместно снижается в основном по причинам перелова и ухудшения условий для нереста.
Рыбец является ценным объектом промысла и его хозяйственное использование может принести значительную прибыль в сложившихся условиях дефицита рыбных продуктов (по последним данным в РФ потребление рыбы на душу населения составляет около 10 – 13 кг в год, в то время как в других странах достигает до 80 кг).
Сенгилеевское водохранилище расположено в 15 км западнее г. Ставрополя (рис. 12). Оно образовано в 1952 – 1958 гг. путем заполнения бывшего соленого (7,3 г/л) Сенгилеевского озера кубанской водой. В настоящее время водохранилище при максимальном проектном наполнении имеет площадь 4200 га, глубину до 38 м, в среднем 15 м, и незначительную (0,5 г/л) соленость воды. Ежегодно с апреля по ноябрь водохранилище наполняется водой до проектной отметки, а в декабре – марте его уровень срабатывается на 8—10 м, в результате чего вся мелководная зона – место нереста фитофильных рыб – осушается.
Биомасса планктона составляет 12,3 г/м3, бентоса – 18,0 г/м3. В состав ихтиофауны водохранилища входит 18 видов рыб: сазан, лещ, синец, густера, судак, рыбец, шемая, форель, подуст, усач, серебряный карась, голавль окунь, ерш, уклея, шиповка, пескарь, игла. Одни из них обитали в бывшем озере, другие зашли в водохранилище с кубанской водой, третьи занесены с целью акклиматизации из других водоемов.

Рис. 12. Схема Сенгилеевского водохранилища
В 1952 г. из рыбопитомника «Горячий ключ» Краснодарского края в Сенгилеевское водохранилище было завезено 5,6 тыс. личинок рыбца. Опыт интродукции дал положительные результаты, поэтому по рекомендации кафедры зоологии Ставропольского сельскохозяйственного института в водохранилище повторно было завезено из Аксайско-Донского осетрового завода 334 тыс. молоди рыбца средним весом 0,5 г в 1964 и 25 тыс. средним весом 1,0 г в 1965 г. Последующие наблюдения показали, что акклиматизация проходного кубанского рыбца в Сенгилеевском водохранилище прошла успешно, рыбец хорошо растет и нормально размножается в условиях замкнутого пресного водоема, т. е. что он превратился в жилую форму (Астанин, 1968).
Изучив биологические особенности рыбца, особенности водоёма вселения, используя опыт предыдущих акклиматизационных работ, было решено в качестве метода акклиматизации использовать активный метод, подразумевающий вмешательство человека в процесс акклиматизации переселенца в водоёме путём проведения рыбоводно-мелиоративных и охранных мероприятий.
Как способ интродукции рыбца, был выбран способ рыбоводного освоения – исходный материал в виде икры или личинок, или взрослых особей заготовляют в р. Кубань и доставляют на рыбоводное предприятие. Далее инкубируют доставленную икру и выращивают молодь до жизнестойкой стадии, которую выпускают в новые для ее вида водоемы. Заселять данные водоемы можно также молодью, полученной от маточного стада переселенца, созданного на рыбоводном предприятии. Выпуск жизнестойкой молоди рыбца в водоём осуществляют в июле – августе. Промысловый возврат рыбца составляет 0,5 – 5 %.
Завершающим этапом процесса акклиматизации является подведение итогов о проделанной работе и оценка результатов.
План акклиматизационных работ.
1.Выбор водоёма вселения, его изучение.
2.Отлов производителей на путях миграции осенью.
3.Получение жизнестойкой молоди на рыбоводном предприятии «Горячий ключ».
4.Транспортировка молоди к местам выпуска.
5.Выпуск посадочного материала.
6.Обработка результатов, выявление стадии натурализации (через 8-10 лет).
Выводы
1. Рыбец обитает в озёрах преимущественно в бассейнах Чёрного, Азовского и Каспийского морей. Половая зрелость наступает на 3 – 5 году жизни при длине тела 15 – 18 см., и массе 125 – 150 г. Нерест с мая по июль при температуре 18 – 24 0С. Плодовитость от 11,4 до 136,4 тыс. икринок. Промысловый вид, но численность не велика.
2. Рыбец относится к теплолюбивым рыбам, которые нерестятся в основном в конце весны – начале лета при температуре воды не выше 15 – 250С. Икра развивается в таком же температурном диапазоне. Содержание кислорода в воде ниже 6 мл/л приводит к удушью. По отношению солености относится к эвригалинным рыбам. Наиболее благоприятной для жизни является нейтральная или слабощелочная реакция воды (рН 7,0-7,5).
3. Для стимулирования половых продуктов у европейской ряпушки применяют экологический метод, при этом производителей выдерживают в искусственных стационарных садках 12 месяцев.
4. Рыбец относится к животноядным рыбам, предпочитая крупные формы ракообразных. При выращивании личинок и ранних мальков в искусственных условиях используют рецепт корма РК-С3М, а при выращивании мальков и сеголеток создан рецепт АК-1КЭ, отличающийся высокой эффективностью.
5. Транспортировка икры, личинок, молоди и взрослых особей можно осуществлять различными методами. В настоящее время для транспортировки икры применяются контейнеры различных конструкций. Наиболее широкое применение получил пенопластовый контейнер. Для перевозки предличинок, личинок и молоди весьма удобной тарой является полиэтиленовые пакеты. А при перевозке взрослых особей часто используют брезентовые чаны вместимостью 2-3 м3 и металлические фляги вместимостью около 40 литров.
6. Целесообразность акклиматизации рыбца заключается в его промысловой и пищевой ценности. В связи с этим необходимо расширять его естественный ареал обитания путем вселения в другие водоемы.
Список использованной литературы
1) Астанин Л.П., Саманеева Л.И. – Сравнительное изучение плодовитости рыбца и шемаи, акклиматизированных в Сенгелеевском водохранилище. – Вопр. ихтиологии, 1968, т.8, вып.4, с. 646 – 653
2) Атлас пресноводных рыб России: В 2-х / Под. Ред. Ю. С. Решетникова. – М.: Наука, 2002. - 379.
3) Берлянд Т.Б. – Нерестилища и условия размножения Каспийского рыбца. – труды ВНИРО, т.24,1953. – 317с.
4) Берг А.С. – Рыбы пресных вод России. – М.: Лёгкая и пищевая промышленность, 1923. – 535с.
5) Иванов А.П. – Рыбоводство в естественных водоёмах. – М.: Агропромиздат, 1988. – 367с.
6) Карпевич А.Ф. – Теория и практика акклиматизации водных организмов. – М.: Лёгкая и пищевая промышленность, 1981. – 297с.
7) Касымов А.Г. – Каспийское море. – Л.: Гидрометеоиздат, 1987. – 154с.
8) Моисеев П.А., Азизова Н.А., Куранова И.И. – Ихтиология. – М.: Лёгкая и пищевая промышленность,1981. – 384с.
9) Никоноров С. И., Ананьев В. И. Комплексные меры по масштабному и ускоренному развитию аквакультуры в Российской Федерации // «Рыбное хозяйство», 2007. №3. С. 33 – 35
10) Павлов Д.С. Биологические основы управления поведением рыб в потоке воды,- М: Наука. 1979.120с.
11) Печников А. С., Шурухин А. С. Развитие рыбного хозяйства во внутренних водоёмах России // «Рыбное хозяйство», 2006. №5. С. 34 – 36.
12) Понамарева Е. Н. Бахарева А. А. Методы транспортировки икры, спермы, личинок, молоди и производителей рыб. Астрахань 2002
13) Привезенцев Ю. А. Рыбоводство: учебник для студентов вузов/ Ю. А. Привезенцев, В. А. Власов. – М.: Мир, 2004. – 456 с.
14) Рыбец (Комплексные исследования в нескольких точках ареала). под. ред. Рыжов А. П. – Вильнюс: изд-во «Моклас»,1987. – 189с
15) Скляров В.Я., Гамыгин Е.А., Рыжков Л.П. Справочник по кормлению рыб. - М.: Лег. и пищ. Пром-сть, 1984. – 120
16) Справочник рыбовода по исскуственному разведению промысловых рыб/ под ред. д. б. н. проф. Н. И. Кожина–М. Пищевая пром. 1971–208 с.
17) Черномашенцев А.И., Мильштейн В.В. – Рыбоводство. – М.: Лёгкая и пищевая промышленность, 1983. – 272с.