
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Рефераты по косметологии
Рефераты по криминалистике
Рефераты по криминологии
Рефераты по науке и технике
Рефераты по кулинарии
Рефераты по культурологии
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Рефераты по косметологии
Рефераты по криминалистике
Рефераты по криминологии
Рефераты по науке и технике
Рефераты по кулинарии
Рефераты по культурологии
Курсовая работа: Гинецей растений
Курсовая работа: Гинецей растений
Содержание
Общая характеристика гинецея. Значение и строение
Происхождение и эволюция гинецея
Плацентация и её типы
Завязь и её типы
Строение и типы семязачатка
Общая характеристика гинецея. Значение и строение
Гинецей - совокупность плодолистиков (карпелей). На плодолистике развивается семязачаток, внутри которого формируется зародышевый мешок - женский гаметофит, поэтому гинецей - это женский орган полового размножения. Обычно плодолистики срастаются краями, образуя замкнутое вместилище - пестик. Из пестика после оплодотворения развивается плод.
Для репродукции растений гинецей имеет первенствующее значение, ибо он, после оплодотворения, разрастается в плод с семенами. Сообразно этому его значению и разные части его имеют различную степень важности для растений. Важнейшая его часть есть семязачаток, превращающийся в семя, Поэтому гинецей без семязачатков лишается своего биологического значения и считается недоразвитым.
Нормальных растений без семяпочек нет, имеются однако же растения, гинецей которых представлен только семяпочкой. Затем по важности следует рыльце, то есть орган, принимающий пыльцу. Настоящего рыльца нет только у голосемянных, где оно заменено однако же рыхлой тканью, развивающейся при отверстии семяпочки (микропилярном). Менее важен столбик, которого вовсе нет у многих растений (многие лютиковые, барбарис и пр.). Настоящей завязи не имеется только у голосемянных.
Пестик состоит из завязи, столбика и рыльца. Те же части различают и у карпелей. Завязь несет семязачатки, и выполняет функцию влажной камеры, предохраняет семязачатки от высыхания и поедания насекомыми и отчасти от колебаний температуры. Из семязачатков после оплодотворения развиваются семена. Рыльце пестика воспринимает пыльцу, поэтому часто поверхность рыльца неровная, бугорчатая и покрыта липкой жидкостью. Столбик приподнимает рыльце вверх. Иногда столбика нет, и образуются сидячие рыльца.
Положение завязи в цветке зависит от степени срастания ее с цветоложем и околоцветником. Различают верхнюю, нижнюю и полунижнюю завязь.
Верхняя завязь свободно располагается на цветоложе, не срастаясь с другими частями цветка. В этом случае части околоцветника располагаются под завязью, поэтому эти цветки еще называют подпестичными. Когда завязь полностью срастается с цветоложем и основаниями лепестков, тычинок, образуется нижняя завязь. Цветолистики прикрепляются к цветоложу над завязью, образуется надпестичный цветок. У полунижней завязи с цветоложем срастается только ее нижняя часть.
Предполагается, что нижняя завязь более надежно защищена от внешних неблагоприятных условий.
Гинецей у разных растений чрезвычайно разнообразен не только по своему составу, числу частей и указанным выше обстоятельствам, но ещё по форме и относительным размерам своих частей. У голосеменных растений он состоит из двух или нескольких семяпочек, незамкнутых в завязь; вместо завязи у них часто чешуевидный листок, при основании которого они и сидят (сосны, пихты, ели и пр.). У остальных цветковых всегда имеется более или менее замкнутая завязь, вследствие чего их и называют покрытосеменными. Кроме того, имеются такие растения, у которых весь цветок состоит из одного только гинецея даже без всякого при нём покрова (ивы).
В настоящее время разделяют три вида гинецеев:
1) Апокарпный гинецей - состоит из множества самостоятельных плодолистиков, отличается краевой плацентацией.
2) Монокарпный гинецей - состоит из единственного пестика с краевой плацентацией.
3) Ценокарпный гинецей - состоит из нескольких сросшихся пестиков. Выделяют три разновидности ценокарпного геницея:
а) Синкарпный - несколько сросшихся стенками плодолистиков, плацентация центрально-угловая.
б) Паракарпный - то же, но стенки плодолистиков не сохранились. Плацентация постенная.
в) Лизикарпный - отличается от паракарпного колончатой (центральной) плацентацией.
Гинецей бывает простым и сложным. Сложный, или апокарпный гинецей состоит из нескольких несросшихся плодолистиков (малина, лютик).
Простой гинецей может содержать либо один плодолистик, тогда его называют монокарпным, либо один пестик из множества сросшихся плодолистиков - это ценокарпный гинецей. Различают три подтипа ценокарпных гинецеев по способу срастания плодолистиков: синкарпный, паракарпный и лизикарпный. У синкарпного гинецея плодолистики срастаются боковыми поверхностями, при этом образуется многогнездная завязь, а семязачатки находятся ближе к центру этой завязи (яблоня, хлопчатник). Паракарпный гинецей образован плодолистиками сросшимися краями, поэтому завязь одногнездная (виноград, крыжовник). Лизикарпный гинецей развивается из синкарпного, но при этом растворяются стенки внутри завязи и семязачатки оказываются расположенными на колонке в центре завязи (гвоздичные).
Структура семязачатка до оплодотворения включает в себя семяножку (фуникулус), плаценту, два интегумента и нуцеллус. Плацента это место прикрепления семяножки к завязи. Внутренняя часть семязачатка - нуцеллус покрыта двумя покровами - интегументами, внешним и внутренним. Интегументы на верхушке семязачатка не смыкаются, образуя микроскопическое отверстие - пыльцевход, или микропиле. Часть нуцеллуса, обращенного к микропиле, называют микропилярным полюсом, а противоположную сторону - халазой, или халазальным полюсом.
После того, как из клетки нуцеллуса образуется мегаспора, она прорастает в семязачатке в зародышевый мешок, занимающий большую его часть, а нуцеллус остается в виде небольшого пристенного слоя.
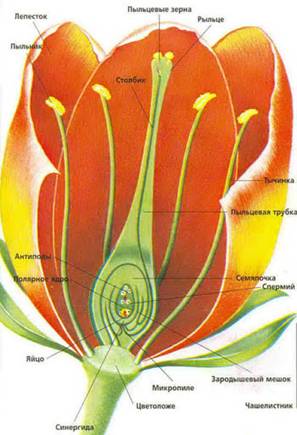
Рис.1 Гинецей
Происхождение и эволюция гинецея
В процессе эволюции гинецея плодолистики постепенно срастаются между собой и из апокарпного гинецея возникает ценокарпный (от греч. kainos - общий) В ценокарпном гинецее отдельные столбики (стилодии) могут оставаться свободными или же срастаются, образуя один общий столбик (сложный столбик). Ценокарпный гинецей бывает трех типов: синкарпный, паракарпный и лизикарпный.
Синкарпным (от греч. syn - вместе) называется гинецей из различного числа замкнутых плодолистиков, сросшихся между собой боковыми частями. Он дву - многогнездный и характеризуется тем, что семязачатки расположены вдоль швов замкнутых плодолистиков, т.е. по углам, образованным брюшными их частями (так называемая угловая плацентация). Синкарпный гинецей обычно происходит из апокарпного гинецея с циклическим (круговым) расположением плодолистиков, но в некоторых случаях он явился производным и от спирального гинецея. Хорошим примером синкарпного гинецея могут служить лилия и тюльпан. На первых порах эволюции синкарпного гинецея срастаются только завязи плодолистиков, а столбики (стилодии) остаются свободными. Но постепенно процессе срастания захватывает также столбики, которые в конце концов срастаются в один сложный столбик, заканчивающийся рыльцевой головкой, что можно видеть у вересковых или у большинства однодольных, в том числе у лилии.
Гораздо чаще синкарпного гинецея встречается паракарпный гинецей (от греч. para - возле,). Для паракарпного гинецея характерна разомкнутость отдельных плодолистиков при сохранении связи краевых участков соседних плодолистиков. В отличие от синкарпного гинецея паракарпный гинецей морфологически одногнездный и характеризуется так называемой постенной или париетальной плацентацией (от лат. parietalis - стенной, рис. 19). Общеизвестные примеры паракарпного гинецея - мак, огурец, тыква. Во многих случаях паракарпный гинецей произошел из синкарпного. Вероятно, это было связано с тем, что с возникновением синкарпного гинецея необходимость в замкнутости каждого отдельного плодолистика во многих по крайней мере случаях становится, в сущности, уже излишней. Поэтому неудивительно, что паракарпный гинецей возник независимо и параллельно в самых разных линиях эволюции цветковых растений и характеризует многие семейства и целые порядки. Конструктивно паракарпный гинецей более экономичен, чем синкарпный: при относительно меньшей затрате строительного материала обеспечивается вполне эффективная защита и снабжение семязачатков и развивающегося зародыша. Кроме того, пыльцевые трубки имеют более широкий доступ к семязачаткам, чем в многогнездной завязи синкарпного гинецея.
Во многих других случаях паракарпный гинецей произошел скорее всего непосредственно из апокарпного. Таково, вероятнее всего, происхождение паракарпного гинецея некоторых африканских родов семейства анноновых, большинство представителей которого характеризуется апокарпным гинецеем, или же происхождение паракарпного гинецея семейства канелловых. Таково же, по всем данным, происхождение паракарпного гинецея представителей семейств савруровых и кактусовых, всего порядка фиалковых и целого ряда происшедших от него порядков, а также семейства водокрасовых среди однодольных.
В паракарпном гинецее обычно происходит разрастание плацент, которые часто даже ветвятся. В очень многих случаях эти разросшиеся плаценты сталкиваются в центре завязи, срастаются между собой и полость одногнездного гинецея разделяется на камеры, представляющие собой ложные гнезда. Разросшиеся плаценты хорошо выражены у некоторых тыквенных, например у огурца и арбуза.
В некоторых линиях развития двудольных, например у портулаковых и первоцветных, из синкарпного гинецея возник лизикарпный (от греч. lysis - развязывание, освобождение, растворение). Это одногнездный гинецей, у которого плацентация не париетальная, как у гинецея паракарпного типа, а так называемая свободная центральная, или колончатая. Одногнездность лизикарпного гинецея возникает в процессе эволюции (или в онтогенезе) в результате исчезновения перегородок (боковых стенок) синкарпной завязи. При этом сросшиеся краевые части не разрушаются и не расходятся, и на них по-прежнему продолжают сидеть семязачатки. Таким образом, эти сохраняющиеся краевые части плодолистиков вместе с плацентами превращаются в центральную колонку, возвышающуюся в полости завязи.
Кроме этих трех основных типов ценокарпного гинецея, различают еще так называемый псевдомономерный гинецей. Это очень редуцированный вариант синкарпного или паракарпного гинецея, в котором фертилен и полностью развит лишь один плодолистик. Стерильные плодолистики в псевдомономерном гинецее часто настолько редуцированы, что их присутствие удается обнаружить только в результате специального изучения их анатомического строения и развития. Типичный псевдомономерный гинецей можно наблюдать у ильма, шелковицы, крапивы, конопли и родственных им растений.
У многих цветковых растений, в том числе у примитивных групп, гинецей еще свободный, не сросшийся с окружающими его частями цветка. Но во многих линиях эволюции цветковых растений совершенно независимо и параллельно произошло большее или меньшее срастание гинецея (как правило, ценокарпного) с окружающими частями цветка и в результате возникли так называемая нижняя завязь. Этот процесс происходил постепенно, и имеются все промежуточные формы между верхней и нижней завязью. Поэтому в морфологии цветка, кроме терминов "верхняя" и "нижняя" завязь, существует также термин "полунижняя" завязь. Происхождение нижней завязи является одним из трудных вопросов эволюционной морфологии растений, и ему посвящена большая литература. Для решения этого вопроса большое значение имело сравнительно-анатомическое изучение цветка, особенно его проводящей системы. Значение проводящей системы в решении различного рода морфологических вопросов основано на ее сравнительно большем постоянстве (консерватизме). Данные по анатомии проводящей системы цветков с нижней завязью приводят к выводу, что в подавляющем большинстве случаев нижняя завязь произошла в результате срастания с завязью оснований чашелистиков, лепестков и тычиночных нитей. Прирастание к завязи тесно облегающей ее цветочной трубки вполне естественно и биологически оправдано. Разные стадии этого прирастания цветочной трубки к завязи можно проследить, например, в семействе вересковых. Однако, как показали те же сравнительно-анатомические исследования, происхождение нижней завязи может быть и иное. У некоторых семейств, таких, как аизооновые, кактусовые, санталовые и родственные им семейства, а также у некоторых других завязь обрастает не цветочная трубка, а цветоложе. Ход проводящих пучков ясно показывает, что завязь как бы погрузилась в чашеобразное углубление цветоложа и срослась с ним. Однако этот второй путь образования нижней завязи наблюдается лишь у очень небольшого числа семейств.
Плацентация и её типы
Плацента́ция - способ расположения плацент и семяпочек (семязачатков) на плодолистиках в завязях цветковых растений. Различия в плацентации имеют важное значение в познании эволюции цветковых растений.
Расположение плацент, как и размещение семяпочек у разных групп цветковых растений неодинаково и тесно связано с тем или иным типом гинецея. Различают два основных типа плацентации: сутуральную, или краевую и ламинальную, или поверхностную.
В апокарпных гинецеях (в том числе и мономерных) отчётливо выражена сутуральная, или краевая плацентация. В этом случае семязачатки располагаются по краям плодолистиков в два ряда вдоль брюшных швов.
В ценокарпных гинецеях (синкарпном, паракарпном и лизикарпном) встречаются разные типы сутуральной плацентации. Это связано с особенностями срастания плодолистиков.
Для синкарпного гинецея характерно срастание кондупликатных или инволютных плодолистиков. Они срастаются боковыми частями, образуя перегородки, разделяющие сформировавшиеся гнёзда завязи, число которых соответствует числу сросшихся плодолистиков. Брюшные швы плодолистиков, несущих плаценты, оказываются в центре завязи. Такую плацентацию называют центрально-краевой, или центрально-угловой (лилейные, колокольчиковые, бурачниковые).
В паракарпном гинецее плодолистики срастаются не боковыми поверхностями, а краями, образуя одногнёздную завязь. Плацентация в этом типе гинецея постенная, или париентальная: постенно-краевая, если плаценты расположены по краям плодолистиков и постенно-спинная, если они приурочены к средним жилкам плодолистиков. Она характерна для видов многих семейств и порядков (капустные, маковые, фиалковые, ивовые, орхидные и др.).
Лизикарпный гинецей также имеет одногнёздную завязь, но плаценты в ней находятся в поднимающейся со дна завязи колонке. Такую плацентацию называют колончатой, или осевой (иногда - свободной центральной). Колончатая плацентация характерна для первоцветных, гвоздичноцветных.
Для примитивных покрытосеменных была характерна ламинальная плацентация, которая встречается и у некоторых современных цветковых (магнолия, лотос, сусак, роголистник, кувшинка). Этот тип плацентации характеризуется прикреплением семязачатков не вдоль брюшного шва, а по всей внутренней поверхности завязи или в определённых её местах.
Место прикрепления семязачатков в завязи называется плацентой. Плацента обычно имеет вид небольшого вздутия, выроста или выступа, образованного тканями завязи.
В зависимости от особенностей прикрепления семязачатков к стенке завязи различают несколько типов плацентации (рис.2). Расположение семязачатков может быть пристенным, или париентальным, когда семязачатки располагаются внутри завязи по ее внешним стенкам или в их выпячиваниях. В случае когда семязачатки находятся на центральной колонке завязи, расчлененной на гнезда по числу плодолистиков, плацентация называется осевой или аксиальной. При свободной центральной плацентации семяпочки развиваются на свободной центральной колонке, не связанной перегородками со стенкой завязи. В некоторых случаях единственный семязачаток находится в самом основании одногнездной завязи. Такой тип плацентации называется базальным. Существуют и другие типы плацентации, но встречаются они сравнительно редко. Различие в плацентации имеет большое значение в систематике цветковых растений.
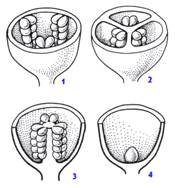
Рис.2.: 1 - постенная, или париетальная, 2 - осевая, или аксиальная, 3 - свободная центральная, 4 - базальная.
Завязь и её типы
За́вязь - термин морфологии растений; замкнутое полое вместилище, нижняя вздутая часть пестика обоеполого или женского цветка. Завязь содержит надежно защищённые семяпочки. После оплодотворения завязь превращается в плод, внутри которого находятся семена, развившиеся из семязачатков [1].
Завязь выполняет функцию влажной камеры, предохраняющей семязачатки от высыхания, колебания температуры и поедания их насекомыми [2].
Завязь и рыльце пестика, которое служит для улавливания пыльцы, соединяются столбиком (если в цветке несколько пестиков, их верхние суженные части называют стилодиями) [3].
Завязь может быть одно - или многогнёздной (в последнем случае она разделена перегородками на гнёзда; иногда гнёзда разделяются ложными перегородками).
По типу расположения в цветке завязи называются:
Верхняя (свободная) завязь - прикрепляется основанием к цветоложу, не срастаясь ни с какими частями цветка (в этом случае цветок называют подпестичным или околопестичным).
Нижняя завязь - находится под цветоложем, остальные части цветка прикрепляются у её вершины (в этом случае цветок называют надпестичным).
Полунижняя завязь - срастается с другими частями цветка, гипантием или цветоложем, но не у самого верха, верхушка её остается свободной (в этом случае цветок называют полунадпестичным).
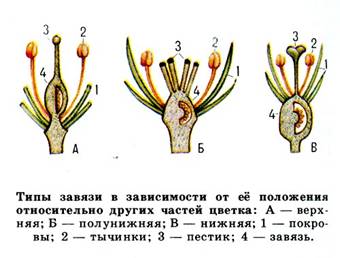
Рис.3. Типы завязей
Строение и типы семязачатка
Семязачаток или семяпочка, многоклеточное образование у семенных растений, из которого развивается семя. Основные части С. - нуцеллус, интегумент (или интегументы) и семяножка. Нуцеллус возникает в типичных случаях в виде бугорка из клеток плаценты мегаспорофилла (плодолистика). Интегумент закладывается в виде кольцевого валика в основании нуцеллуса и обрастает развивающийся нуцеллус, оставляя над его вершиной узкий канал - микропиле, или пыльцевход, под которым у большинства голосеменных находится пыльцевая камера. Семяножка (фуникулус) соединяет. с плацентой. базальную часть, от которой отходит семяножка, называемая халазой.
В завязи пестика располагаются небольшие образования - семязачатки. Их число колеблется от одного (пшеница, слива) до нескольких миллионов (у архидных). Функции семязачатка - мегаспорогенез (создание мегаспор) и мегагаметогенез (формирование женского гаметофита, процесс оплодотворения). Оплодотворенный семязачаток развивается в семя. Плацента - место прикрепления семязачатка к плодолистику.
Части семязачатка:
нуцеллус, ядро семязачатка;
фуникулюс, семяножка, с помощью которой семязачаток крепится к плаценте;
интегументы, покровы семязачатка, образующие на верхушке нуцеллуса канал;
микропиле, пыльцевход;
халаза, базальная часть семязачатка, где сливаются нуцеллус и интегументы;
рубчик - место присоединения семязачатка к семяножке.
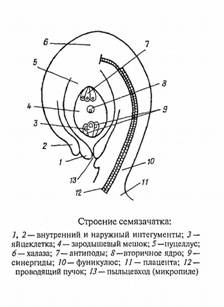
Рис.4 Строение семязачатка
Типы семязачатка:
ортотропный - прямой, фуникулюс и микропиле расположены на противоположных концах оси семязачатка (гречишные, ореховые);
анатропный (обратный) - нуцеллус повернут по отношению к прямой оси семязачатка на 180, вследствие чего микропиле и фуникулюс расположены рядом (Покрытосеменные)
гемитропный (полуповернутый) - семязачаток повернут на 90, вследствие чего микропиле и нуцеллус расположены по отошению к фуникулюсу под углом 90 (первоцветные, норичные).
кампилотропный (односторонне изогнутый) - нуцеллус изогнут односторонне микропилярным концом, соответсвенно микропиле и фуникулюс расположены рядом (бобовые, мальвовые)
амфитропный (двусторонне изогнутый) - нуцеллус изогнут двусторонне в виде подковы, при этом микропиле и фуникулюс располагаются рядом (тутовые, ладанниковые).
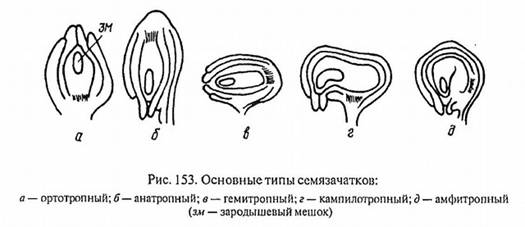
Рис.5 Основные типы семязачатков
6. Развитие семязачатка
Мегаспорогенез протекает в женской репродуктивной сфере - в гинецее. Морфологически гинецей представлен пестиком (или пестиками). В состав пестика входят: рыльце, столбик и завязь. Внутри завязи содержатся семязачатки (один или несколько). Внутреннее содержимое семязачатка представляет собой нуцеллус. Покровы семязачатка образованы двойным или одиночным интегументом. В нуцеллусе семязачатка имеется одна археспориальная клетка (2n), способная делиться путем мейоза (у ив и некоторых других растений археспорий многоклеточный). В результате мейоза из археспориальной клетки (материнской клетки мегаспор) образуется четыре гаплоидные мегаспоры (n). Вскоре три из них отмирают, а одна увеличивается в размерах и трижды делится путем митозов. В результате образуется восьмиядерный зародышевый мешок (женский гаметофит). Три ядра вместе с прилегающей цитоплазмой образуют клетки-антиподы, два ядра - одно центральное диплоидное ядро; два ядра - две клетки-синергиды; одно ядро становится ядром яйцеклетки.
Когда к семязачатку приближается пыльцевая трубка, он это заранее "чувствует" и готовится ее встретить. Клетки-спутницы начинают выделять слизистые вещества. Тем временем пыльцевая трубка растет, преодолевая сопротивление клеток стенок завязи. Наконец, она достигает микропиле. Происходит "драматический" процесс: пыльцевая трубка протыкает (и при этом убивает) одну из клеток-спутниц. Оба спермия покидают пыльцевую трубку. Участь вегетативной клетки пыльцевого зерна незавидна, она вскоре погибнет. Наблюдать этот процесс очень трудно, но еще труднее понять, что происходит при оплодотворении.
В августе 1898 года, когда еще не было известно о двойном оплодотворении, в Киеве состоялся десятый Съезд русских естествоиспытателей и врачей. Профессор Сергей Гаврилович Навашин сделал на нем важное сообщение: оба спермия, содержащиеся в пыльцевом зерне, необходимы для нормального развития семян двух видов из семейства Лилейные (Liliaceae): лилии (Lilium martagon) и рябчика (Fritillaria tenella). Почему Навашин выбрал именно эти растения? Наверное, потому, что спермии и зародышевый мешок у них крупные, их легко рассматривать в микроскоп. Навашев впервые заявил, что в процессе оплодотворения участвуют два спермия, сливающиеся с двумя (!) клетками зародышевого мешка. Но предоставим слово самому автору открытия.
Каждый раз, когда пыльцевая трубка наблюдалась в соприкосновении с зародышевым мешком, оба мужские половые ядра также наблюдались в содержимом зародышевого мешка. Мужские ядра лежат сначала близко друг от друга...
Мужские ядра затем отделяются друг от друга, причем одно проникает к яйцеклетке, а другое тесно прикладывается к одному из еще не слившихся в это время полярных ядер, именно к сестринскому ядру яйцеклетки...
В то время, как мужское ядро все более и более тесно прилегает к ядру яйца, полярное ядро, копулирующее с другим мужским ядром, направляется навстречу к другому полярному ядру, с которым встречается в середине зародышевого мешка...
Только после прохождения профазы деления ядра сливаются..."
Чтобы объяснение было понятнее, Сергей Гаврилович сделал к рисунки, очень понравившиеся столичным ботаникам. Они отвезли их, чтобы показать на заседании Петербургской Академии.
Почему же этому выступлению ученые придали такое большое значение? Считалось, что оплодотворение у растений происходит точно так же как у животных. Один спермий и одна яйцеклетка должны давать зиготу, из которой развивается новое растение. Не то чтобы ботаники не видели двух спермиев, движущихся в пыльцевой трубке к зародышевому мешку. Считалось, что это - какая-то ненормальность, "типичное" растение должно иметь не два, а один спермий, "типичное" оплодотворение должно быть одинаковым и у растении и у животных. Оказалось, что эти представления были неправильными. Оплодотворение цветковых растений совершенно не похоже на оплодотворение не только животных, но и других растений. Процесс оплодотворения с помощью двух спермиев получил название двойного оплодотворения. Сразу же после открытия Навашина ученые бросились к своим препаратам. Оказалось, что многие уже видели двойное оплодотворение, но считали его уродливым процессом, якобы не дающим нормальных семян. Ученые стали присылать Сергею Гавриловичу поздравления. А один ученый даже подарил свои старые препараты, на которых было хорошо видно двойное оплодотворение.
Итак, С.Г. Навашин открыл удивительное явление. Зачем происходит оплодотворение яйцеклетки было понятно: чтобы получилась зигота, а из нее - новое растение. Но зачем же оплодотворять центральную клетку зародышевого мешка? Оказалось что именно из этой клетки развивается питательная ткань семени - эндосперм. Навашин предположил, что без мужского ядра эндосперм образоваться не может. Как же проверить это предположение? Ведь из пыльцевой трудки невозможно вытащить один из спермиев, не нарушая процесс оплодотворения.
Сергей Гаврилович решил исследовать растения, у которых нет эндосперма. Вдруг у них нарушено оплодотворение центральной клетки? Такие растения отыскались в семействе Орхидных (Orchidaceae). Семена орхидных очень мелкие, и даже прорости самостоятельно не могут (а прорастают только при помощи грибов базидиомицетов, образуя микоризу). В пыльцевой трубке орхидных Навашин увидел два спермия. Один из них оплодотворял яйцеклетку, а второй "пробовал" оплодотворить центральную клетку с двумя полярными ядрами. Но в центральной клетке ядра не сливались друг с другом! Процесс оплодотворения был нарушен и, естественно, эндосперм не мог образоваться. В дальнейших работах ученому удалось показать, что у подсолнечника (Helianthus annuus) и некоторых других растений оплодотворение - двойное. После того, как Навашин нашел двойное оплодотворение у самых разных растений, он сделал вывод, что двойное оплодотворение свойственно всем цветковым растениям.
Взглянем на строение семязачатка после двойного оплодотворения. Клетки обоих его покровов как имели два набора хромосом, так их и сохранили. Оба набора в этих клетках принадлежат материнскому растению. Нуцеллус также несет два материнских наборов хромосом. В зародышевом мешке синергиды погибли, а у антиподов по одному набору хромосом. Эти наборы - тоже материнские. Зигота, образовавшаяся слиянием яйцеклетки и спермия, несет два набора хромосом: один от отцовского, а другой от материнского растения. Самое интересное, что центральная клетка имеет три набора хромосом: один от отцовского растения, и два - от материнского.
После двойного оплодотворения начинается несколько процессов: первичное ядро эндосперма делится, образуя эндосперм, зигота развивается в зародыш, интегументы превращаются в семенную кожуру, а стенка завязи и связанные с ней структуры формируют плод. На ранних стадиях развития последовательность клеточных делений у зародышей двудольных и однодольных сходна; в обоих случаях формируются сферические тела. Позже появляется различие: зародыш двудольных имеет две семядоли, а однодольных - только одну. В семенах некоторых групп покрытосеменных нуцеллус развивается в запасную ткань, называемую перистермом. Некоторые семена содержат и эндосперм и перистерм, так у свеклы (Belta). Однако у многих двудольных и некоторых однодольных вся запасная ткань или большая ее часть поглощается развивающимся зародышем еще до того, как семя впадет в состояние покоя (у гороха, у бобов и др.). Зародыши в таких семенах обычно развивают заполненные питательными веществами мясистые семядоли. Основные питательные вещества, запасаемые в семенах, - углеводы, белки, липиды. Семена голосеменных и покрытосеменных различаются происхождением этих веществ. У первых их дает женский гаметофит, у вторых - эндосперм, не являющийся ни гаметофитной, ни спорофитной тканью.
Развитие семяпочки в семя сопровождается превращением завязи (а иногда и других частей растения) в плод. В ходе этого стенка завязи (околоплодник, или перикарарпий) часто утолщается и дифференцируется на отдельные слои - внешний экзокарпий (внеплодник), средний мезокарпий (межплодник) и внутренний эндокарпий (внутреплодник), - обычно лучше заметные в мясистых, чем в сухих, плодах.
Так происходит развитие гаметофита, оплодотворение и формирование семени у "типичного" цветкового растения, однако мир растений очень разнообразен и у множества покрытосеменных развитие проходит иначе, однако не менее интересно и удивительно. Все эти способы развития помогают каждому растению поддерживать свою стратегию в борьбе за существование; преимущество двойного оплодотворения не ограничивается тем, что растения формируют питательные запасы в семязачатке только при условии оплодотворения (не "растрачивая" силы на формирование питательных запасов в семязачатке, который никогда не будет оплодотворен, как это часто происходит у голосеменных), эти различия в развитии, являющиеся следствием приспособленности растений к окружающим их условиям, и создают то удивительное многообразие форм морфологического и физиологического строения, которое мы и наблюдаем у цветковых.
7. Зародышевый мешок и его развитие
Женский гаметофит цветковых растений обычно называется зародышевым мешком. Возникновение этого названия восходит к тем временам, когда еще не была ясна морфологическая природа чрезвычайно своеобразного женского гаметофита цветковых растений. В настоящее время оба термина употребляют как синонимы.
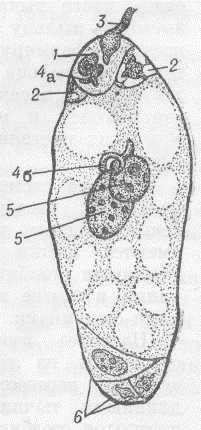
Рис.6. Зародышевый мешок (женский заросток) покрытосемянного (цветкового) растения и двойное оплодотворение: 1 - яйцеклетка; 2 - клетки-помощницы; 3 - вскрывшаяся пыльцевая трубка. Один из спермиев (4а) сливается с яйцеклеткой; 5 - полярные ядра, сливающиеся со вторым спермием (46); 6 - три клетки в нижней части зародышевого мешка (антиподы).
У большинства цветковых растений (вероятно, не менее чем у 80%) начало женскому гаметофиту дает одна из четырех потенциальных мегаспор тетрады путем трех последовательных митотических делений. Этот тип образования женского гаметофита называется моноспорическим. Его развитие происходит следующим образом. В результате первого деления ядра функционирующей мегаспоры образуются два ядра, которые расходятся к полюсам (микропилярному и халазальному) сильно удлиняющейся мегаспоры, а между ними образуется крупная вакуоль. Далее каждое из этих двух ядер делится синхронно еще два раза и в результате у каждого полюса образуются по четыре ядра. Это восьмиядерная стадия развития женского гаметофита. От каждой из двух полярных четверок входит по одному ядру в центральную часть зародышевого мешка. Эти так называемые полярные ядра сближаются в центральной части и сливаются или сразу же, или только перед оплодотворением, а иногда и вовсе не сливаются. В результате слияния полярных ядер образуется ядро центральной клетки женского гаметофита, называемое часто вторичным ядром зародышевого мешка.
В дальнейшем (в процессе последнего митоза) возникают клеточные перегородки и свободноядерная стадия развития женского гаметофита переходит в клеточную стадию. У микропилярного полюса образуются три клетки яйцевого аппарата, у противоположного (халазального) - три клетки, называемые антиподами, а между этими двумя группами клеток - центральная клетка (содержащая два свободных полярных ядра или одно вторичное ядро). Три клетки яйцевого аппарата дифференцируются на одну яйцеклетку и две клетки, называемые синергидами (от греч. synergos - вместе действующие) или вспомогательными клетками. Таково строение так называемого нормального моноспорического женского гаметофита.
Нормальный моноспорический женский гаметофит не производит впечатления окончательно отработанной модели, в которой экономичность достигла такого же предела, как в случае мужского гаметофита. То, что предел здесь не достигнут, доказывается следующим. В процессе эволюции этого основного для цветковых растений типа женского гаметофита возникли зародышевые мешки других типов, образуемые двумя или даже четырьмя мегаспорами. Произошло сокращение митотических делений до двух или одного, а также изменилось распределение ядер. Разные сочетания этих изменений обусловили возникновение нескольких типов женского гаметофита, которые отличаются как числом ядер (4, 8, 16), числом клеточных групп и полярных ядер (1, 2, 4, 7-14), так и числом клеток в группах (например, яйцевой аппарат может состоять из 1, 2, 3, 5 и 7 клеток), а также другими признаками.
Основной особенностью моноспорического типа образования женского гаметофита является последовательное заложение клеточных перегородок при мейотических делениях и образование обособленных мегаспор. Совершенно иной характер имеет мегаспорогенез, предшествующий образованию биспорического (двухспорового) и тетраспорического (четырехспорового) гаметофитов. Изменения в мегаспорогенезе заключаются в подавлении заложения клеточной перегородки при мейотическом делении, вследствие чего вместо тетрад мегаспор возникают неклеточные образования, называемые ценоцитами (от греч. koinos - общий и kytos - сосуд, клетка), которые и развиваются далее в женские гаметофиты. Здесь возможны два варианта.
В первом случае образование клеточных перегородок подавляется только при втором делении мейоза. Поэтому оба ядра, вошедшие в состав жизнеспособной клетки диады, принимают участие в образовании женского гаметофита. Двухъядерная диада - это ценоцит, продукт слияния (вернее, неразделения) двух потенциальных мегаспор. Поскольку каждый такой ценоцит соответствует по происхождению паре необособившихся мегаспор, его рассматривают как двухспоровый ценоцит, а развивающийся из него женский гаметофит - как биспорический (двуспоровый). Биспорический женский гаметофит наблюдается у лука, пролески (Scilla), ландыша, амариллисовых, частуховых и некоторых других растений. Развитие такого женского гаметофита происходит более сокращенным путем и для образования восьмиядерной стадии требуется всего лишь два митотических деления.
Наконец, у некоторых растений, таких, как, пеперомия, ильм, майник, рябчик, тюльпан, лилия, гусиный лук, плюмбаго, адокса и другие, образование клеточных перегородок подавляется при обоих мейотических делениях. Вся материнская клетка мегаспор превращается здесь в один четырехспоровый ценоцит, или тетраценоспору. Четырехспоровый ценоцит соответствует четырем неразделившимся мегаспорам, почему женские гаметофиты этого типа называются тетраспорическими (четырехспоровыми). Для образования восьмиядерной стадии женского гаметофита требуется лишь одно митотическое деление ядер четырехспорового ценоцита.
Согласно И.Д. Романову, в отличие от моноспорического женского гаметофита, би - и тетраспорические называются ценоспорическими.
Женский гаметофит цветковых растений произошел из более примитивного архегониального женского гаметофита далеких голосеменных предков путем выпадения последних стадий развития, ведущих к формированию архегониев. А у ценоспорических типов происходит, как мы видели, выпадение также начальных стадий. Но в то время как эволюция мужского гаметофита достигла кульминационной стадии и в его развитии полностью выпали все возможные начальные и конечные стадии, с женским гаметофитом эволюционный процесс как бы продолжает экспериментировать. Отсюда значительное число моделей женского гаметофита и путей его формирования, на чем здесь нет, однако, возможности останавливаться.
Литература
1. http://ru. wikipedia.org/wiki
2. И.И. Андреева, Л.С. Родман "Ботаника"/ Москва "КолосС" 2002
3. П. Рэйвн, Р. Эверт, С. Айкхорн "Современная ботаника"/ Москва "Мир" 1990
4. Магешвари П., Эмбриология покрытосеменных, пер. с англ., М., 1954; Тахтаджян А.Л., Высшие растения, т.1, М. - Л., 1956;
5. Поддубная-Арнольди В.А., Общая эмбриология покрытосеменных растений, М., 1964;
6. Савченко М.И., Морфология семяпочки покрытосеменных растений, Л., 1973.