
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Рефераты по косметологии
Рефераты по криминалистике
Рефераты по криминологии
Рефераты по науке и технике
Рефераты по кулинарии
Рефераты по культурологии
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Рефераты по косметологии
Рефераты по криминалистике
Рефераты по криминологии
Рефераты по науке и технике
Рефераты по кулинарии
Рефераты по культурологии
Курсовая работа: Вивчення мохоподібних району польової практики - околиць сіл Копили та Розсошенці
Курсовая работа: Вивчення мохоподібних району польової практики - околиць сіл Копили та Розсошенці
Зміст
Вступ
Розділ 1. Характеристика природніх умов м. полтави та його околиць
Розділ 2. Матеріали і методика дослідження
Розділ 3. Мохоподібні як особлива лінія еволюції вищих рослин
Розділ 4. Мохоподібні лісових ценозів околиць сіл розсошенці та копили
4.1 Морфолого-біологічні особливості зібраних мохів
4.2 Еколого-ценотичні особливості виявлених мохів
Розділ 6. Господарське значення мохоподібних і їхня роль у природі
Висновки
Список використаної літератури
Вступ
Мохоподібні є однією з найменш досліджених груп рослинного світу. Тому вивчення особливостей їх анатомічної і морфологічної будови, еколого - ценотичних властивостей та участі в утворенні рослинного покриву набуває все більшої актуальності. Вони є досить великою, близько 25 тис. видів, групою рослинного світу. Це представники вищих, або пагонових рослин. Мохоподібні найбільш примітивні автотрофні вищі рослини. Від більшості інших рослин вони відрізняються відсутністю коренів і деякими мікроскопічними особливостями.
Мохоподібні поділяються на три відділи: антоцеротові (Аnthocerotophyta), печіночники (Нераtіcophyta) і мохи (Bryophyta). Усі три відділи виникли на Землі дуже давно, близько 300 мільйонів років тому, і з тих пір розвивалися незалежно один від іншого, а тому поряд із загальними ознаками, що вказують на походження їх від одного предка, вони володіють і рядом специфічних, властивих тільки їм особливостей.

Рис.1. Маршанція.
1) чоловічий таллом, 2) жіночий таллом.
Тому метою нашої роботи і було вивчення мохоподібних району польової практики - околиць сіл Копили та Розсошенці, виявлення їх морфолого-біологічних та еколого-ценотичних особливостей.
Завдання нашої роботи наступні:
1) збір та гербаризація мохоподібних;
2) визначення зібраних видів;
3) виявлення морфолого-біологічних та еколого-ценотичних особливостей зібраних видів;
4) визначення частоти трапляння мохів;
5) встановлення участі виявлених видів в утворенні біоугруповань.
Розділ 1. Характеристика природніх умов м. полтави та його околиць
Територiя Полтавської області (площа 28, 8 тис. га) розташована в межах Днiпровсько-Донецької западини, якiй в сучасному рельєфi вiдповiдає Придніпровська низовина, представлена на пiвднi Приднiпровською терасою, в центральнiй та пiвнiчнiй частинах - Полтавською рiвниною. Рельєф територii рiвнинно-хвилястий, розчленований долинами рiчок на вододiльні плато. Глибина залягання грунтових вод коливається в значних межах вiдповідно до елементів рельєфу: на заплавi 2-3 м; на боровій терасі - 3,5-12 м; на лесовiй терасі - 4 - 10 м; на плато - 22 - 24 м.; в балкових системах, що перерізують плато - 6 - 7 м. (Бєлосельська, 1968). Грунтові води характеризуються значною мiнералізацією, яка зростає в пiвденнiй частині регіону. На територii областi досить розвинена гідрологiчна мережа, яку складають річки (головнi з них лiвi притоки Днiпра - Ворскла, Суда, Псел), болота, озера, штучнi водойми. Щiльнiсть рiчкової мережі складає 0,15 км/км (Клименко, 1958). Згiдно з клiматичним районуванням Полтавщина розташована в межах Антлантико-Континентальної областi (Щербань, 1984) i характеризується найменшим зволоженням серед лiсостепової зони України. Клiмат помiрно континентальний. Особливiстю клiмату регiону є поступове зменшення в напрямку на схiд i пiвдень кількості опадiв (з 600 до 490 мм, вiдповiдно i показникiв випаровування) та пiдвищення в лiтнiй перiод температур, а також тривала посуха та суховії в південній та пiвденно-схiднiй частинi, що обумовлене межуванням iз континентальною степовою областю. Середньорiчна температура повiтря склдлає 5,7-6,6°С. Середня температура сiчня мiнус 6 - 8°С. Тривалiсть зимового перiоду - 122 днi. Середня температура влiтку (липня) - 19-20°С. Тривалiсть лiтнього перiоду - 120 днiв (Бабиченко и др., 1967).
Грунтовий покрив регiону в цілому досить строкатий, але переважають чорноземи глибокi малогумуснi на пiдвищених частинах стародавнiх терас Днiпра. У пiвнiчнiй частинi поширенi глибокi малогумуснi вилуженi чорноземи, якi чергуються на зниженнях з лучно-чорноземним ґрунтами. На терасах рiчок Сули, Псла, Ворскли поширенi чорноземно-лучнi солонцюватi грунти, серед яких специфiку рослинного покриву визначає наявнiсть солонців, солончакiв та солончакуватих грунтiв. Для пiвнiчних районів характерне содове засолення, для центральної та пiвденної частин - хлоридне та сульфатне. Чималi площi займають торфовi грунти, переважно в пiвнiчно-захiднiй частинi, зокрема, в долинi Сули, де приуроченi до рiчкових заплав та знижень на межирiччях.
Лiсовi грунти - чорноземи опiдзоленi та темно-сiрi опiдзоленi подекуди в комплексi з сiрими i свiтло-сiрими - займають переважно розчленованi правобережжя рiчок Сули, Псла, Ворскли. На борових терасах рiчок поширенi дерновослабкопiдзолистi, пiщанi та глинисто-піщані грунти. Із негативних явищ у регіоні слід вiдмiтити площинну i яружну ерозiю, утворення зсувiв, усiдання, оглеєння i засолення грунтiв. Найпоширенішими на території Полтавщини є кленово-липово-дубовi лiси, якi репрезентують типові для Лівобережного Лiсостепу широколистянi лiси. Найбiльшi масиви кленово-липово-дубових лiсiв пов’язанi з крутим i найбiльш розчленованим правобережжям рiчок Сули, Псла, Ворскли та iх приток. Трав’яний покрив широколистяних лiсiв флористично небагатий i складається з типових неморальних видiв. В умовах Лiсостепу зростає вплив екологiчних факторiв, що обумовлюють ксеро - та галофiтизацiю рослинного покриву. Це вiдбивається i на розподiлi деревної рослинностi в заплавах регiону дослiджень (Андриенко, 1982). Соснові ліси областi характеризуються незначною фiтоценотичною та флористичною рiзноманiтнiстю, що пояснюється подiбними умовами їх формування на борових терасах рiчок iз збiдненими супiщаними грунтами та штучним походженням. Найпоширенiшi угруповання представленi середньовiковими сосновими насадженнями, що займають пiдвищені та вирiвнянi дiлянки борових терас (М’якушко, 1972). Цi флористично збiдненi ценози характеризуються невисоким проективним покритгям травостою iз значною участю лишайникiв та мохiв. Дубово-соснові лiси займають бiльш родючi грунти борових терас - глинисті піски з суглинковими прошарками або легкі супіски. Характерною особливiстю рослинного покриву Лiвобережного Приднiпров’я в межах лiсостепової зони України є зростаюча галофiтизацiя, пов’язана з особливостями геоморфологii та ґрунтового покриву. На терасах Днiпра та його лiвобережних приток поширенi солонцi та солончаки (особливо в пониззях). Солончакуватi мулувато-глеєвi та дерново-глеєві слабозасоленi грунти приуроченi до притерасних та центральних частин заплав вздовж усiєї течiї рiчок. Рослинний покрив регiону незважаючи на всебiчний антропогенний вплив, характеризується значним рiзноманiттям. Найвищi показники ценотичного i флористичного багатства притаманнi степовим i лучним екосистемам, дещо меншi - лісовими та гiдрофiльним.
Флористична рiзноманiтнiсть Полтавщини також має вагомi кiлькiснi показники: квiткових нараховується понад 1500 видiв, голонасiнних - 3 види в природних умовах (понад 100 видiв i рiзновидностей в культурi), папоротевидних - 16, хвощв - 9, пiлаунiв - 3, мохiв - 159, лишайникiв - 161 вид (Байрак, 1997; Байрак та iн., 1998).
Природнi компоненти територiї (геологiчна будова, рельєф, клiмат, вода, грунти, рослинний i тваринний світ), перебуваючи мiж собою в складному довготривалому зв’язку, утворюють рiзноманiтнi, закономiрно побудованi природно-територiальнi комплекси або ландшафти.
Сучасний рельєф Полтавщини сформувався пiд впливом ендогенних (внутрiшнiх) та екзогенних (зовнiшнiх) сил Землi, переважно протягом кайнозойської ери, шо почалася 60 - 70 млн. рокiв тому. Основнi риси сучасного рельєфу були закладені в неогеновий перiод цієї ери пiсля поступового вiдступу на пiвдень Харкiвського моря і формування річкової сiтки.
Бiльша частина областi розташована в Приднiпровськiй низовині, крайня пiвденно-західна (правобережна) частина - в межах Приднiпровської височини, крайня пiвнiчно-схiдна частина - частково на вiдрогах Середньоросiйської височини. Поверхня областi являє собою слабкохвилясту некруто похилу до Днiпра рiвнину, розчленовану долинами рiчок Сули, Псла, Ворскли, Орелi та їх приток.
За характером рельєфу лiвобережну частину областi можна подiлити на три частини: пiвденно-захiдну, центральну, пiвнiчно-схiдну. Пiвденно-захiдна, що тягнеться вздовж Днiпра, найнижча, малорозчленована. Тут немає ярiв та балок, характерних для iнших частин областi. Помiтним уступом по лiнії Кобеляки - Хорол - Пирятин вiддiляється вiд пiвденно-захiдної центральна частина, бiльш розчленована рiчковими долинами, ярами та балками. Пiвнiчносхiдна частина - найбiльш підвищена i найбiльш розчленована, в зв’язку з цим має складний горбистий характер. В цілому рівнинна територія області в зв’язку з її розчленованiстю рiчковими долинами, сiткою ярiв i балок, дислокацiями має складний рельєф. Нерідко на рiвнiй мiсцевостi пiдносяться досить високi горби. За походженням рельєф областi переважно ерозiйний, тобто вироблений талими водами льодовикiв антропогенового перiоду, річками та тимчасовими водотоками. Основними i найбiльш поширеними формами рельєфу є вододiльнi плато, рiчковi долини, балки, яри. З неерозiйних форм рельєфу мають мiсце степовi блюдця, льодовиковi та солянокупольні дослокації. Територiя Полтавської областi належить до помiрного клiматичного поясу, крайньої пiвденної частини Атлантико-континентальної помiрно вологої помiрно теплої клiматичної областi. Характер (тип) клiмату взагалi обумовлюється комплексом чннникiв (факторiв), основу яких складають:
1) сонячне випромiнювання (радiацiя);
2) характер циркуляції повiтряних мас;
3) близькiсть великих водних мас (океанiв, морiв);
4) висота територiї над рiвнем моря;
5) тип (характер) пiдстилаючої поверхнi. З урахуванням характеру цих чинникiв клiмат областi може бути визначений як помiрно континентальний.
Сонячне випромінювання. Тепло на поверхню Землi надходить у виглядi прямої і розсiяної сонячної радiації. Максимум прямої сонячної радiацiї в областi припадає на липень, мiнiмум - на грудень. В середньому за рiк тривалiсть сонячного сяяння (ТСС) становить 1894 - 2021 годину, що складає 43-48% від можливого.
Температура повітря. Середня рiчна температура повітря в межах області змiнюється від +6,5° С на пiвнiчному сходi до +8, З°С на пiвденному заходi. Найхолоднiший зимовий мiсяць - сiчень. Середня мiсячна температура сiчня становить 5,3° - 7,2°С морозу. Найнижча температура повiтря настає при надходженнi в наші широти арктичного повiтря. Абсолютний мiнiмум температури повiтря на територii областi - 38°С. Найтеплiший мiсяць року - липень. Середня температура липня 19,8°С - 21,6°С тепла. Середній максимум температури повiтря за рiк становить 10,8°С - 13°С тепла, в сiчнi - 2,4°С - 4,4°С морозу, в липнi 25,6°С - 28°С тепла. На Полтавщинi теплий перiод триває з 14.03 - 22.03 по 18.11 - 25.11 (тривалість 240 - 255 днiв).
Особливостi атмосферноi циркуляцій. Циркуляцiя повiтряних мас є базовим фактором формування погодних та клiматичних умов. Територiя Полтавської областi знаходиться на межi впливу Атлантичного океану i Центральної Азії, а також пiвнiчних i пiвденних повiтряних мас. В середньому в рiчному циклi на територii областi переважає захiдна форма циркуляцii повiтряних мас (40%), потiм схiдна (34%), яка характеризується впливом зi сходу i пiвнiчного сходу Сибiрського антициклону, переважно взимку. Кожна з повiтряних мас має свої фiзичнi характеристики. При зiткненнi повiтряних мас відбувається не тільки їх взаємний обмiн теплом i вологою, але, що особливо важливо, утворення атмосферних завихрень - циклонiв i антициклонiв. З циклонами пов’язана нестiйка хмарна погода з опадами; з антициклонами - стiйка безхмарна сонячна погода. Циклонiчний тип погоди панує на територiї областi в середньому 135 днiв на рiк. Протягом року територiя областi перебуває пiд впливом 35 - 36 антициклонiв. Антициклонiчний тип погоди панує в областi протягом 230 днiв на рiк. Атмосфернi опади. Важливою погодною i клiматичною характеристикою є вiдносна вологiсть повітря, що показує мiру насичення повiтря водяною парою і дозволяє судити про можливiсть чи неможливiсть опадiв (при вiдноснiй вологостi повiтря 100% починається конденсацiя водяної парни i угворення опадiв). Середня вiдносна вологiсть повiтря в областi становить 71%; найменша (58%) в серпні; найбiльша (88%) - в сiчнi. Однiєю з найбiльш важливих характеристик клімату є атмосфернi опади. Для Полтавської областi характерний континентальний тип рiчного розподiлу опадiв з максимумом влiтку і мiнiмумом зимою. Середня рiчна кiлькiсть опадiв на територiї областi коливається в межах 450 - 570 мм, що вiдповiдає 4500 - 5700 т/га. За холодний перiод року (листопад - березень) опадiв випадає в середньому 130 - 170мм, а за теплий (квiтень - жовтень) 320 - 400 мм. В окремi роки рiчна сума опадiв може вiдхилитись вiд середньорічної. Найчастiше сильнi дощi та зливи бувають при перемiщеннi пiвденно-захiдних та пiвденних циклонiв. Опади з кiлькiстю 50 мм i бiльше за добу приносять збитки [15].
Розділ 2. Матеріали і методика дослідження
Збір видів, гербаризація та визначення мохоподібних проводились із дотриманням загальних правил збору та згідно існуючих вимог (А.С. Лазаренко, 1955, А.Ф. Бачурина, Л.Я. Партина, 1979)
Виконану нами роботу можна розділити на 3 основні етапи:
польові збори та вивчення місцевостей зростання мохів;
обробка та визначення зібраних матеріалів;
обробка літературних джерел з даної теми.
При зборі зразка ми користувались ножем та польовими пакетами. До кожного додавалась чорнова етикетка, на якій ми вказували місцезнаходження, по можливості назву роду.
При визначенні мохоподібних ми користувались мікроскопом МВN - 3, бінокуляром, чашкою Петрі, препарувальними голками, предметними і покривними скельцями.
При ідентифікації зразків здійснювалася таким чином:
Спочатку відмічається колір, щільність дернини, а також колір та наявність спорогонів.
Частина зразку відмочується у воді, після чого відмічається: довжина стебла, форма та колір листків, наявність спорогонів, форма коробочки, кришечки.
За допомогою мікроскопа досліджуємо анатомічну будову листка:
жилкування;
форму листкової пластинки (край, середину, основу);
будову перистома коробочки;
характер ніжки.
Особливу увагу звертали на диференціацію клітин основи листка, а саме: колір та розмір.
При визначенні зразків ми використовували такі визначники:
Лазаренко А.С. Определитель лиственных мхов Украины.
Мельничук В.М. Определитель лиственных мхов средней полосы и Юга Европейской части СССР.
Гарибова Л.В. Водоросли, лишайники и мохообразные СССР.
Розділ 3. Мохоподібні як особлива лінія еволюції вищих рослин
На відміну від нижчих рослин (водоростей), тіло більшості мохоподібних представлено пагоном, що складається зі стебла і листів; тільки в їх частини воно являє собою слань або талом.
Від нижчих рослин мохоподібні відрізняються також численними мікроскопічними особливостями, у тому числі наявністю своєрідно улаштованих гаметангіїв (статевих органів): чоловічих - антеридіїв і жіночих - архегоніїв.
Відмінна ознака мохоподібних - правильне чергування в нормальному циклі розвитку рослини двох різних за своєю морфологією поколінь. Одне з поколінь називається гаметофітом (рослина, що виробляє статеві елементи - гамети), інше - спорофітом (рослина, що виробляє елементи безстатевого розмноження - спори). Якщо антеридій й архегонії розташовуються на одному гаметофіті, то такі рослини називаються однодомними. Якщо на одній рослині (чоловічій) розташовуються антеридії, а на іншому (жіночій) - архегонії, то такі види називаються дводомними. Є і багатодомні мохоподібні, у яких антеридії й архегонії можуть розташовуватися на одному і на різних рослинах того ж виду.
При наявності краплинно - рідкого середовища сперматозоїд досягає яйцеклітини і запліднює її. З зиготи виростає спорофіт, що у мохоподібних називається спорогоном, який складається зі стопи, ніжки та коробочки. За допомогою ніжки спорогоній висмоктує з гаметофіту воду з мінеральними солями й органічними речовинами. У листяних та антоцеротових мохів так як має хлорофіл він частково живиться самостійно. У коробочці спорогонія утворюється споровий мішок, або спорангій, в якому в результаті мейозу утворюються спори. Дозріла коробочка розкривається, спори попадають у зовнішнє середовище. При наявності сприятливих умов вони проростають і дають початок протонемі, яка має вигляд багатоклітинної нитки, пластинки, кулястого тіла й ін., а потім уже виростає гаметофор - власне сланевий або листостебловий гаметофіт, що несе гаметангії, у яких знову виникають сперматозоїди і яйцеклітини. У такий спосіб відбувається чергування поколінь у життєвому циклі мохоподібних. Детально цикл розвитку розглянемо на прикладі зозулиного льону.
Зозулин льон - Polytrіchum commune.
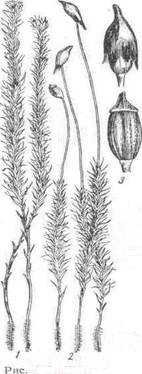
Рис.2.
1) чоловіча рослина,
2) жіноча рослина зі спорогоніями,
3) спорогонії.
Зозулин льон (Polytrіchum commune) є характерним представником листяних мохів. Його тіло розчленовано на тонке, округле, червонуватого кольору стебло і вузькі, зелені листочки. Корені відсутні, їх заміняють добре розвинуті ризоїди. У порівнянні з іншими видами мохів зозулин льон досягає у висоту 20-40 см.
Розмножується він спорами. У цього виду добре виражена зміна поколінь. Це дводомна рослина.
Чоловічі екземпляри зозулиного льону мають характерне росташування листків на верхівці стебел. Тут утворюються більші листки, сидять вони значно густіше у виді розеточки і мають червонувате зафарбування. По такому розташуванню листочків легко знайти чоловічі екземпляри. Антеридії формуються на розширеній верхній частини стебла. Вони мають трохи подовжену форму і у них розвиваються сперматозоїди з двома джгутиками. Архегонії мають колбоподібну форму, розташовані на верхівці стебла жіночої рослини, що, на відміну від чоловічої, не закінчується розеткою з червоних листочків. Запліднення відбувається ранньої весни, коли низькі місця, на яких виростають мохи, заливаються водою. Один зі сперматозоїдів проникає до яйцеклітини через ослизнений канал шийки архегонію і запліднює її. Із заплідненої яйцеклітини виростає спорофіт у вигляді довгої тонкої стеблинки, що закінчується коробочкою складної будови. Коробочка спорогона має подовжений із загостреним кінцем ковпачок. Зовні він подібний із зозулею, звідки і назва цього моху. Ковпачок (каліптра) - це верхня змінена частина архегонію. Під ковпачком знаходиться кришечка коробочки. Всередині коробочки є центральний стрижень - колонка, навколо неї міститься споровий мішок, у якому розвиваються спори. Спочатку вони з'єднані в тетради, тобто по чотири разом. Перед дозріванням тетради розпадаються на окремі спори. У коробочки спочатку відпадає ковпачок, потім кришечка. Край урночки коробочки закінчується зубчиками (перистом), у суху погоду вони відгинаються назовні і тим самим відкривають вихід зрілим спорам.
Спора, випадаючи на землю, при наявності достатньої кількості вологи проростає, утворюючи протонему, або передросток. Протонема складається з тонких гіллястих ниток, заповнених хлорофілом. Вона утворює верхівкову бруньку, з якої утворюються дорослі рослини зозулиного льону, причому одні протонеми утворюють тільки чоловічі, а інші тільки жіночі рослини. Хоча зовнішнього розходження між спорами немає, фізіологічно вони різняться. Після запліднення яйцеклітина розростається в безстатеве покоління у вигляді спорогона що росте на жіночому гаметофіті. У зозулиного льону гаметофіт за своїми розмірами переважає над спорофітом. Це рослина багаторічна. Після звільнення від сперматозоїдів чоловічі екземпляри не гинуть; вони продовжують рости і на наступний рік на верхівці їх знову утворюються антеридії. Не гинуть і жіночі екземпляри, після розсіювання спор спорогон відпадає, а рослини продовжують рости, і на наступну весну на верхівці стеблинки знову утворяться архегонії. [9]
Істотною відмінною рисою вищих рослин, до яких відносять і мохоподібних, від нижчих є наземний спосіб життя. У результаті тривалої еволюції вищі рослини вийшли з води і пристосувалися до життя в нових своєрідних наземних умовах. У процесі пристосування до наземного способу життя у них виробилося багато різних ознак і властивостей, відбулася диференціація органів і ускладнилась анатомічна будова, що сприяло нормальному способу життя у різноманітних умовах суші. Однією із гіпотез утворення мохоподібних є походження їх від водоростеподібних предків, Ймовірніше всього це була група зелених водоростей - археохарофіт (Рановський, 1979). При проростанні спор у мохів розвивається гілляста зелена нитка - протонема, що нагадує тіло нитчастих водоростей. Статевий процес у мохів здійснюється тільки у водному середовищі. Це вказує на споріднення мохів і водоростей.
Зозулин льон широко розповсюджений у вологих лісах, на луках, інколи на полях лісової зони. Низькорослі прямостоячі рослини нерідко утворюють щільний травостій. На стеблі багато листків, але знизу вони відсутні. Нижня частина стебла часто переходить у кореневище, що росте під землею горизонтально. Кореневище густо покриті ризоїдами, розростаючись, дають початок новим рослинам.
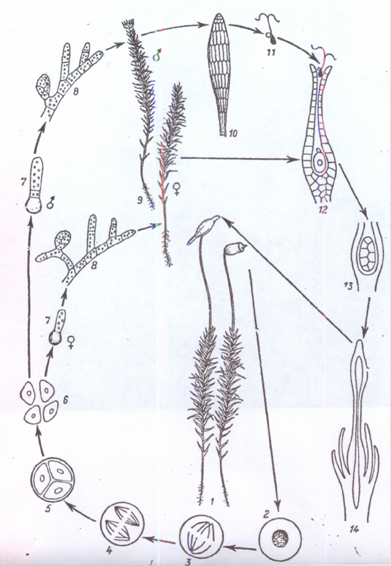
Схема циклу розвитку зозулиного льону (Polytrіchum commune):
1 - дорослі рослини зі зрілими спорогонами; 2 - спорогенна клітина; 3, 4, 5 - утворення спор (редукційний поділ); 6 - спори; 7, 8 - утворення чоловічої та жіночої протонем; 9 - чоловічій та жіночій гаметофіти; 10 - антеридії; 11 - сперматозоїд; 12 - архегоній з яйцеклітиною та сперматозоїдом; 13, 14 - розвиток спорогона.
Розділ 4. Мохоподібні лісових ценозів околиць сіл розсошенці та копили
В результаті наших досліджень встановлено, що в лісових масивах околиць сіл Розсошенці та Копили виявлено 19 видів мохоподібних. Вони належать до 8 родин, до 12 родів і 15 видів.
Таблиця 1.
Видовий склад зібраних мохоподібних
|
РОДИНА Polytrichaceae Dicranaceae Funariaceae Bryaceae Mniaceae Brachytheciасeae Hypnaceae Endodontaceae |
ВИД Polytrichum juniperinum Hedw. Atrichum undulatum (Hedw) Beauv. Dicranum scoparium Hedw. Dicranum polysetum Sw. Ceratodon purpureus (Hedw) Brid. Funaria hygrometrica Hedw. Pohlia nutans (Hedw) Lindb. Mnium cuspidatum Hedw. Brachythecium oldipodium B. S. G. Eurhynchium hians (Hedw) Saegev. Platygyrium repens (Brid) Bryol eur. Pylaisia polyantha (Hedw) B. S. G. Hypnum pallescens (Hedw) Beauv. Hypnum cupressiforme Hedw. Pleurozium schreberi (Brid) Mitt. |
4.1 Морфолого-біологічні особливості зібраних мохів
Відділ Мохи (Bryophyta)
Клас Брієві мохи (Bryopsida)
Підклас Брієві (Brydae)
Прядок Політрихові (Polytrichales)
Родина Політрихові (Polytrichaceae)
Рід, вид Зозулин льон ялівцевий (Polytrichym juniperum Hedw)
Дернинки сизувато-зелені, густі. Стебла до 10см, прямостоячі. Листки при основі лінійно-ланцетні; часто із злегка загнутою всередину чи відігнутою назовні верхівкою. Край цілісний, широко завернений всередину. Жилка зверху на спинці зубчаста. Повздовжні пластинки листка з гладенькою, пляшкоподібною, краєвою клітиною. Внутрішні листки по краю широкі; білошкірі з довгою віссю.
Коробочка видовжена, призматична, з різко виділеною дископодібною шийкою. Зубців перистому 64. Ковпачок найчастіше довший за коробочку, волосистий.
По всій території України на відкритих, сухих місцях, пісках, в сухих хвойних чи листяних лісах, на скелях, камінні, на рівнині і в горах. [3] ; [10]
Цей мох був знайдений нами в околицях с. Копили, в сосновому лісі на згарищі.
Рід, вид Атріх хвилястий (Аtrichum undulatum (Hedw) Beauv) Одно - або дводомний. Дернинки рихлі, темно - або жовто-зелені. Стебло до 10 см висоти. Сухі листки - скручені і зігнуті всередину, вологі - прямостоячі, хвилясті, 5-10 мм, лінійно - язиковидні, загострені. Жилка закінчується у верхівці листка.
Спорогони зазвичай поодинокі, рідко на верхівці пагону розташовуються 2 (3) спорогона. Ніжка 2-4 см, червонувата або жовтувата. Коробочка зазвичай нахилена, близько 5 мм, червонувато - або темно-бура. Спороносить восени.
Широко поширений по всій лісовій смузі України; зазвичай на порушеному або незадернованому ґрунті в хвойних і змішаних лісах, рідше на луках і покритих гумусом каменях в горах.
Цей мох ми знайшли у листяному лісі.
Порядок Дикранові (Dicranales)
Родина Дикранові (Dicranaceae)
Рід, вид Дикран віничний (Dscranum scoparium Hedw)
Дернинки зелені, бурі, блискучі, пухкі. Стебло 3-5 см. висотою, просте або слабо галузисте. Листки до 2 мм завдовжки, рівномірно відстовбурчені в усі сторони, прямостоячі або серцевидно-односторонні; з широко-ланцетої основи до середини гостро пильчасті, шилоподібні. Жилка досягає верхівки листка, на нижньому боці з 2-3 низькими повздовжніми пластинками. Клітини основи пластинки листка бурі, зверху коротко-прозенхіматичні; внизу лінійні, утворюють вушка, вгорі короткі і шестикутно-ромбічні. Клітини вушок
коричневі, не доходять до жилки.
Коробочка циліндрична, нахилена, темно-коричнева, без жилок, гладенька, кришечка з дуже довгим дзьобиком. На Україні досить поширений вид в соснових лісах, на ґрунті, скелях.
Цей вид ми зустріли в околицях с. Копили в сосновому лісі на ґрунті. Часто.
Вид Дикран багатоніжковий (Dicranaceae polysetum S w) Дернинки м'які, рихлі, зелені або жовтувато-зелені, злегка блискучі. Стебло до 15 см висотою, зазвичай просте, з густою білуватою або бурою ризоїдною повстю. Листя до 1 см довжиною, обернені у всі або в один бік; пластинка яйцевидно-ланцетна, загострена, по краю в нижній половині пильчаста; бурі клітини в кутках основи листа не досягають жилки.
Спорогони по 1-5 з одного перихеція, з жовтими або червонуватими, до 4 см дов., ніжками. Коробочка нахилена або повисла, циліндрична, з маленьким зобиком, в сухому стані - скручена і борозенчаста. Кришечка з довгим дзьобиком. Спороносить влітку і восени.
На ґрунті і гниючих стовбурах в лісах, рідше на скелях і болотах. Характерний широко поширений вид лісової смуги. [8]
Нами вид відмічений у сосновому лісі околиць с. Копили на ґрунті.
Рід, вид Цератодон пурпурний (Ceratodon purpureus (Hedw) Brid) Стебло 1-3 см, прямостояче, розгалуджене. Листки прямостоячі, від яйцевидно-ланцетних до вузьколанцетних, загострені.
Ніжка спорогона до 3 см довжиною, пурпурово-червона, блискуча. Коробочка довгаста, червонувата або бура. Спороносить влітку. Зазвичай на голих ґрунтах, узбіччях доріг, на схилах горбів, згарищах, каменях, а також на гнилій деревині і на солом’яних очеретах та дерев'яних дахах. [8]
Цей вид ми зустріли у сосновому лісі в основі стовбурів дерев.
Родина Funariaceae Рід, вид Фунарія вологомірна (Funaria hygrometrica Hedw) Дернинки від блідо-зелених до темно-зелених. Стебло 1-3 см довжиною. Верхні листки до 4 мм довжиною, бруньковидні скупчені, короткозагострені, суцільнокраї або вгорі пильчасті. Ніжка спорогону 3-5 (8) см довжиною, спочатку зігнута, пізніше пряма, скручена. Коробочка косогрушевидна, повисла або горизонтальна, смугаста, червонувато-бура суха і відкрита. Перистом подвійний. Кришка плоскоопукла. Ковпачок роздутий внизу. Спороносить навесні і літом. Зустрічається повсюди на голому ґрунті, часто на згарищах і поблизу житла. [8]
Цей вид ми знайшли на ґрунті у листяному лісі.
Родина Брієві (Bryaceae) Рід, вид Полія поникла (Pohlia nutans (Hedw) Lindb) Дернинки темно - або жовтувато-зелені (у сухому стані - злегка блискучі, але без бронзового блиску). Стебло до 1-3 (4) см довжиною. Верхівкові листки до 4 мм довжиною, лінійно-ланцетні, загострені, зубчасті, з жилкою, що закінчується під верхівкою листка або виступає з неї.
Ніжка спорогона до 4 см довжиною, червонувата. Коробочка від нахиленої до повислої, від широкоеліпсоїдної до циліндричної, з короткою шийкою. Спороносить влітку. Однодомний.
Дуже мінливий вид. Зростає на ґрунті, гнилій деревині, скелях, в лісах і на болотах. [3]
Цей вид ми зустріли в околицях с. Розсошенці на ґрунті та при основі дерев.
Родина Мнієві (Mniaceae) Рід, вид Мній загострений (Mnium cuspidatum Hedw) Дернинки часто широкі, яскраво-зелені, внизу повстисті. Генеративне стебло прямостояче, 2-5 см довжиною, при основі із стерильними пагонами. Листки до 5 мм довжиною, оберненояйцевидні, загострені, зубчаті у верхівки, облямовані, з жилкою, що закінчується у верхівці листка або під нею. Спорогони поодинокі, на червонувато-жовтих, 1,5-3 см. довжиною, ніжках. Коробочка повисла, еліпсоїдна. Спороносить на початку літа. Однодомний. На затіненому сируватому ґрунті, гниючих пнях і покритих землею скелях, переважно в лісах. [14] Цей вид ми зібрали в листяному лісі околиць с. Розсошенці на ґрунті та в прикореневій частині стовбурів дерев.
Родина Брахітецієві (Brachytheciaceae) Рід, вид Брахітецій коротколистий (Brachythecium oldipodium B. S. G) Дернинки темно - або жовто-зелені, з зеленими матово-блискучими листками та пагонами. Стебло лежаче або висхідне. Стеблові листки з відігнутою верхівкою, низхідні, яйцевидно-серцевидні, звужені в ланцетоподібну верхівку, в верхній частині пильчасті. Жилка закінчується над серединою листка. Клітини листка прозенхіматичні, тонкостінні, в кутах листка багаточисельні. Листки гілочок низхідні, коротко і широко загострені зі скрученою верхівкою, до основи гостропильчасті, жилка на спинці зубчаста. Ніжка спорогона бородавчаста. Коробочка нахилено -горизонтальна, широкояйцевидна, кришечка коротко конічна. [10]
Цей вид ми відмітили на ґрунті та гнилій деревині у сосновому лісі.
Порядок Гіпнові (Hypnales) Родина Гіпнові (Hypnaceae) Рід, вид Платегірій повзучій (Platygyrium repens (Brid) Bryol eur)
Дернинки золотисто - чи коричнево-зелені, блискучі. Стебло повзуче, перисто-розгалужене, з округлими, прямими чи зігнутими гілками. Стеблові листки яйцевидно-ланцетні, цілокраї. Жилки немає або вона помітна і подвійна. Клітини листка лінійні, на верхівці короткі.
Коробочка прямостояча, з короткою шийкою, видовжена або циліндрична, правильна або зігнута, жовтувата або коричнева. Перистом подвійний. Зубці зовнішнього перистома вузьколанцетні.
Цей вид ми зустріли у листяному лісі околиць с. Розсошенці на стовбурі дуба, ясена та ін.
Рід, вид Пілезія багатоквіткова (Pylaisia polyantha (Hedw) B. S. G) Дернинки темно - або жовтувато-зелені. Стебло 2-5 см довжиною. Листки прямостоячі, на кінцях гілок обернені в один бік, яйцевидно-ланцетові. Ніжка спорогона до 2 см довжиною. Коробочка циліндрична, коричнева, прямостояча. Спороносить восени і весною.
На корі дерев, особливо часто на осиці, по стовбурах якої піднімається на декілька метрів, рідше на гнилій деревині і на покритих гумусом каменях.
Зустрічається по всій території України, особливо на півночі, північному - заході. [10]
Цей мох ми зустріли на старому дубі в листяному лісі околиць с. Розсошенці.
Рід, вид Гіпн білуватий (Hypnum pallescens (Hedw) Beauv) Дернинки блідо - або жовтувато-зелені. Стебло 2-4 см довжиною, повзуче, перистогіллясте. Стеблове листки дворядні, довгасто-яйцевидні, з шилоподібною зігнутою верхівкою, зубчаті по всьому краю. Кришечка з косим дзьобиком. Спороносить навесні. Однодомний. На корі дерев в хвойних і змішаних лісах.
Цей вид ми побачили на старому дубі у листяному лісі околиць с. Розсошенці.
Вид Гіпн кипарисовий (Hypnum cupressiforme Hedw)
Дернинки зелені або жовтувато-зелені. Стебло 2-10 см довжиною, неправильно або перистогіллясте, з прямостоячими, дуговидними гілочками. Листки серповидно-гачкуваті, інколи прямі, довголанцетні, шиловиднозагострені з плоскими цілісними або неясно зубчастими краями. Кришечка дзьобовидна. Спороносить навесні. Дводомний. Дуже мінливий вид. На сонячних та затінених скелях у горах, сухих кам’янистих тундрах, в арктичних пустелях, на стовбурах дерев, на гнилій деревині та на ґрунті в лісах. [8]
Цей вид ми побачили у складі знайденого гнізда та у сосновому лісі околиць с. Копили на ґрунті.
Рід, вид Евринхій зіяючий (Eurhynchium hians (Hedw) Saeger)
Дернинки плоскі, жовто-зелені, блискучі. Листки яйцеподібні, звужені в коротку, вузьку верхівку, від основи гостро пилчасті. Жилка тонка, на спинці із шипиком. Клітини прозенхіматичні, у верхівці ромбічні, шестикутні; у кутках квадратні та коротко-прямокутні; листя гілочок яйцевидне, гостропилчасте. Жилка з шипиком. Ніжка спорогону бородавчаста. Коробочка видовжено-яйцеподібна, горбата, бура, кришечка із косим дзьобиком. Поширені по всій території України в тінистих листяних лісах, на затінених скелях, газонах в садах, парках. [10]
Цей мох ми побачили у листяному лісі с. Розсошенці та у складі знайденого гнізда.
Родина Ентодонтові (Endodontaceae)
Рід, вид Плеуроцій Шребера (Pleurozium schreberi (Brid) Mitt) Дводомні мохи, створюють рихлі, від темно-зелених до солом'яно-жовтих, блискучі дерновинки. Стебла 5-15 см. довжиною, висхідні, зазвичай перистогалузисті, з червонуватою корою. Листя ложковидно-увігнуте, складчасте, від широкояйцевидних до продовговато-яйцевидних, тупі або з загострені, з відігнутими вгорі зубчатими краями, короткою подвійною жилкою і оранжевими вушками.
Ніжка до 4 см довжиною, червонувата. Коробочка нахилена або горизонтальна, з короткою шийкою, довгасто-циліндрична, пряма або злегка горбата. Кришечка гостро - або тупоконусовидна. Спороносить восени. На ґрунті, рідше на гнилій деревині і на основах стовбурів дерев, в лісах, в тундрі і на луках.
Цей вид ми знайшли в сосновому лісі околиць с. Копили, на ґрунті.
4.2 Еколого-ценотичні особливості виявлених мохів
Життя мохоподібних, як і життя інших рослин, залежить від багатьох факторів зовнішнього середовища - освітленості, вологості, тепла, складу і руху повітря, хімічного і механічного складу субстрату, на якому вони зростають, прямого і непрямого впливу інших живих організмів. Як і інші зелені рослини, більшість мохоподібних може існувати тільки при достатній кількості світла. Лише використовуючи енергію Сонця, вони здатні створювати необхідні для свого існування органічні речовини з неорганічних. Серед тисяч видів мохоподібних лише деякі види є напівсапрофітами - рослинами, здатними частково харчуватися за рахунок мертвої органічної речовини субстрату і частково створювати органічні речовини з неорганічних за рахунок енергії Сонця. Сапрофітних організмів серед мохоподібних дуже мало. До таких рослин, що живуть тільки за рахунок розкладання мертвої органічної речовини субстрату, відноситься, наприклад, криптоталлус дивний (Cryptothallus mіrabіlіs), таломний підземний печіночник, що живе на лісових сфагнових болотах Західної Європи. [4]
Різні види мохоподібних вимагають для свого існування різної інтенсивності освітлення. Серед них можна виділити і світлолюбні види, що живуть на добре освітлених скелях, і види тіньовитривалі і навіть тіньолюбні, здатні виростати в нішах серед каменів, у печерах, дуплах дерев, під наметом густого темнохвойного лісу, там, де більшість квіткових рослин існувати не може. Яскравий приклад пристосованості мохоподібних до життя в умовах слабкої освітленості - самосвітний мох (схістостега периста). Дуже багато видів мохоподібних можуть успішно рости і розвиватись в досить широких межах освітленості, але при недостатнім освітленні такі рослини нерідко мають світліше зафарбування і більш витягнуті пагони. Залежність мохоподібних від тривалості освітлення ще недостатньо вивчена. У всякому разі багато видів успішно ростуть і спороносять як в умовах неперервного полярного дня, так і в умовах щодобового чергування світла і темряви в помірних широтах (полярну ніч мохоподібні по суті справи проводять у стані "схованого життя" - криптобіозу). Серед виявлених нами мохоподібних 27% видів відносяться до світлолюбів. Це такі, як Polytrichym juniperum, Dicranum polysetum, Dicranum scoparium, Ceratodon purpureus. Тіневитривалі представлені 66% видів: Atrichum undulatum, Pohlia nutans, Mnium cuspidatum, Platygyrium repens, Pylaisia polyantha, Hypnum pallescens, Hypnum cupressiforme, Brachythecium oldipodium, Eurhynchium hians, Funaria hygrometrica, Pleurozium Schreberi
Особливо тісний зв'язок мохоподібних з іншим фактором зовнішнього середовища - вологою. Життя мохоподібних більше, ніж життя інших вищих рослин, залежить від води, що випадає безпосередньо з атмосфери - дощу, туману, роси, а також від водяних парів атмосфери. Це тому, що в циклі розвитку мохоподібних панує гаметофіт, а в циклі розвитку інших вищих рослин, починаючи від плауноподібних і закінчуючи квітковими, переважає спорофіт, що має корені. Сланевий або листостебловий гаметофіт мохоподібних, на якому напів паразитує спорогон, має лише ризоїди, тому він не може, подібно спорофітові плаунів, хвощів і т.п., всмоктувати воду з глибоких горизонтів ґрунту. За допомогою ризоїдів статеве покоління мохоподібних може отримувати воду лише із самого верхнього шару ґрунту, але основну масу води гаметофіт звичайно одержує, всмоктуючи її (у виді краплинно - рідкої води або водяних парів) усією поверхнею свого тіла. Краплинно-рідка вода необхідна мохоподібним також для запліднення. Усі ці морфолого-фізіологічні особливості гаметофіту пояснюють, чому в процесі своєї еволюції мохоподібні освоювали в першу чергу вологі місця, де є достатня кількість доступної для нього води. Саме в таких місцях і понині живе більшість видів бріофітів. [4]
З'явившись на Землі сотні мільйонів років тому, вони змогли пристосуватися до життя в інших місцях. Так, вони змогли освоїти місця з надлишковим зволоженням (ямки і канави з водою, береги струмків і озер, болота), при чому вироблені ними пристосування дозволяють мохоподібним в умовах холодного й помірного клімату нерідко витісняти в таких місцях більшість вищих рослин (звичайно що не виносять одночасно вимокання, низьких температур і недостачі кисню в субстраті). А сфагнові мохи, володіючи своєрідною внутрішньою будовою і хімізмом, навіть накопичують надлишок вологи, виділяючи і підкислюючи її, несприятливо впливаючи на інші рослини. Мохоподібні пристосуватися і до життя на субстратах, без вільної води - на корі дерев, скелях і каменях. Прикріплюючись ризоїдами до такого субстрату, вони одержують вологу лише з атмосфери, а під час відсутності опадів вони можуть переносити висихання, втрачаючи понад 90% води. При випаданні дощу, роси або насиченні повітря парами води рослини швидко всмоктують воду і продовжують рости. Відомі випадки, коли деякі мохоподібні, що пролежали в гербарії кілька років, оживали, будучи змоченими водою.
У цілому серед бріофітів (як і серед інших вищих рослин) можна виділити стосовно води кілька екологічних груп: гідрофіти, гігрофіти, мезофіти, ксерофіти.
Гідрофіти живуть у воді. Вони прикріплюються ризоїдами до стовбурів або гілок дерев, що потонули, або до підвідних каменів (наприклад, фонтиналіс протипожежний - Fontіnalіs аntіpyretіca Hewd) або вільно плавають на поверхні або напівзанурені. Тому розрізняють гідатофіти (рослини повністю занурені у воду) та гідрофіти (рослини, які напівзанурені у воду).
Гігрофіти - рослини дуже зволожених місць (болота, берега річок і струмків і т.п.). Дернинки і коврики гігрофітів, наприклад сфагнум, велику частину року просочені водою. Деякі рослини можуть поводитися і як гідрофіти, і як гігрофіти: наприклад, риччіокарп плаваючий (Rіccіocarpus natans) може плавати на поверхні води або жити на вологому мулистому ґрунті по берегах водойм.
Мезофіти - рослини, що живуть у місцях (часто тінистих) із середніми умовами зволоження (вологі луки, широколистяні і темнохвойні ліси і т.п.).
Ксерофітів, тобто рослин, здатних переносити посуху, не знижуючи сильно життєвої активності, серед мохоподібних нема, і ті з них, що живуть у посушливих, сонячних місцях (скелі, дюни і т.п.), лише умовно можна називати ксерофітами. Здатність таких рослин виростати на сухих місцях у першу чергу забезпечується здатністю їхньої плазми виносити тривале зневоднювання і швидко відновлювати свою структуру при обводнюванні. Ця здатність зв'язана також з різними морфологічними пристосуваннями (зменшення площі листів, наявність волосків з мертвих клітин, заповнених у сухому стані повітрям, скручування або повздовжнє складання листової пластинки або слані при висиханні і т.п.). Між названими групами є перехідні типи. Наприклад, більшість видів, що живуть на корі стовбурів у хвойних і листяних лісах помірного поясу, можна називати ксеромезофітами: під час сильних морозів узимку й влітку, коли відносна вологість повітря дуже низька, ці види повинні вміти справлятися з дефіцитом вологи, в інший же час вони живуть в умовах середньої зволоженості (у насиченому парами води повітрі або на змоченій дощами корі). Серед зібраних нами видів ми виявили такі екогрупи за відношенням до вологи: Мезоксерофіти: 60%; мезофіти: 40%
Тепло в житті мохоподібних відіграє важливу роль, оскільки від температури навколишнього середовища залежить не тільки швидкість випару води рослиною, але також відносна вологість повітря і ґрунту, швидкість обмінних реакцій в організмі. У порівнянні з іншими вищими рослинами серед мохоподібних набагато більше видів, здатних існувати в широких температурних межах, переносити дуже низькі і дуже високі температури. Обумовлюється це в першу чергу тим, що багато мохоподібних легко і без шкоди для себе втрачають воду і саме в безводному стані переносять максимальні і мінімальні температури, перебуваючи фактично у стані криптобіозу. Деякі мохи - ксерофіти в повітряно-сухому стані витримують протягом 30 хв.
Мохоподібні в порівнянні з багатьма іншими вищими рослинами легше переносять несприятливий загально кліматичний вплив ще і тому, що, будучи дуже низькими рослинами, вони фактично живуть в інших температурних умовах, ніж судинні рослини. Ґрунтові мохоподібні живуть в умовах своєрідного, більш м'якого і рівного мікроклімату; температура, вологість повітря, сила вітру і т.п. якого істотно відрізняються від показників погоди. Наприклад, в Антарктиді мохи, що живуть в оазисах на скелях, влітку зазвичай ростуть при позитивних температурах "прискельного" шару повітря, у той час як на рівні 2 м над поверхнею землі температури повітря можуть бути в цей час мінусовими. Перенесенню несприятливих кліматичних умов сприяє також і утворення у мохів дернинок, подушечок, килимів та інших типів "життєвих форм". Всередині подушечки, наприклад, коливання температури і вологості будуть менш різкими, ніж ззовні.
Мохоподібні, подібно лишайникам, реагують на присутність у повітрі шкідливих домішок. Це пов'язано з відсутністю у них особливих покривних тканин і нездатністю більшості мохоподібних щорічно обновляти свій фотосинтезуючий апарат подібно тому, як роблять це квіткові рослини, скидаючи листя восени. Можливо, гострота реакції мохоподібних пов'язана також із невеликою масою їхнього тіла. Але вирішальну роль у їхній чутливості до шкідливих домішок, звичайно, відіграють властивості самої протоплазми, на користь чого може свідчити хоча б той факт, що деякі мохоподібні все-таки здатні жити в межі досить великих міст, атмосфера яких сильно забруднена різними шкідливими домішками. Це так звані стійкі, толерантні види.
Бріофіти практично не зазнають прямого негативного механічного впливу сильних вітрів, оскільки швидкість вітру при поверхні землі або біля якихось перешкод сильно падає. Вітер в основному впливає на рослини непрямо, висушуючи субстрат і збільшуючи транспірацію або ж приносячи опади. На деяких океанічних островах і узбережжях, там, де постійно дмуть вологі вітри з моря, моховий покрив розвинутий набагато пишніше. Позитивну роль відіграє вітер у розселенні мохоподібних - перенесенні спор і навіть шматочків рослин.
Механічний склад субстрату, на якому поселяються мохоподібні, відіграє в їхньому житті меншу роль, ніж у житті інших вищих рослин, оскільки субстрат для них лише місце їхнього прикріплення і джерело для одержання мінеральних речовин. Він не є середовищем, у якій живе значна частина тіла рослини. Проте різні види по-різному відносяться до механічного складу субстрату. У цілому мохоподібні можуть зростати на ґрунті будь-якого механічного складу - на піщаному, супіщаному, глинистому, мулкуватому й ін. Зростають вони і на материнській породі (вапняках, гнейсах, гранітах та ін), щільно прикріплюючись до поверхні скель і каменів ризоїдами, і на субстратах органічного походження (корі і стовбурах дерев, гнилій деревині, екскрементах тварин). Тому за відношенням до субстрату серед мохоподібних виділяють такі групи: епігеї (рослини, що селяться на ґрунті); епіксили (рослини гнилої деревини); епіліти (рослини кам’янистих субстратів); епіфіти (рослини стовбурів дерев); епіфіли (рослини, що селяться на листках дерев). .
Набагато більше значення, ніж механічний склад субстрату, мають для мохоподібних його кислотність і хімічний склад. Деякі види віддають перевагу лужному, вапняковому субстрату, інші можуть жити тільки на дуже кислому субстраті, треті віддають перевагу нейтральному ґрунтові або можуть жити в досить широких межах значення рН. З кислотністю субстрату звичайно пов'язане багатство його важливими для життя рослин мінеральними солями, у першу чергу азотистими сполуками. Сфагнум Магелана, наприклад, задовольняється тими незначними слідами мінеральних солей, які випадають з атмосфери. Та й узагалі дуже багато мохоподібних добре себе почувають на субстратах, що містять невелику кількість живильних мінеральних речовин (піщані і торф’яні ґрунти, вивержені породи і т.п.). Але серед них є види, що віддають перевагу ґрунтам і субстратам, в яких велика кількість поживних речовин, у тому числі сполуки азоту. Наприклад, види роду сплахнум (Splachnum) і багато інших представників родини сплахнових часто поселяються на тваринних залишках, що розклалися.
Для мохоподібних - гідрофітів і гігрофітів середовищем існування є вода; при цьому багато видів успішно ростуть на болотах у стоячій холодній воді з незначним вмістом кисню і мінеральних речовин і з великим вмістом органічних кислот. Корені більшості інших вищих рослин звичайно погано функціонують у такому середовищі. Деякі мохоподібні здатні рости в умовах з великою кількістю мінеральних кислот, теплій і навіть гарячій воді вулканічних джерел. Серед них відсутні види, здатні жити в солоній або солонуватій воді.
Є види бріофітів, що живуть на субстратах з таким вмістом солей і важких металів яке перешкоджає існуванню інших судинних рослин.
Наприклад, види милихоферії (Mіelіchhoferіa), що називаються "мідними мохами", живуть на скелях, з високим вмістом іонів міді, яка у тисячу разів перевищуючи максимальний рівень, припустимий для життя більшості квіткових рослин. Входячи до складу угруповань живих організмів - біогеоценозів, мохоподібні тісно пов'язані з іншими його членами. Вони випробовують на собі вплив інших рослин і тварин і самі впливають на них. Кора дерев і чагарників може служити таким же субстратом для поселення мохоподібних, як і поверхня скель і каменів.
Бріофіти зі свого боку впливають на судинні рослини. Поселяючись на листях дерев вологих тропічних лісів, вони злегка пригнічують їх. У зоні тундри, високо в горах, мохоподібні надають притулок судинним рослинам, стебла і корені яких ховаються в мохових подушках і дернинах від впливу несприятливих кліматичних факторів. Але з другого боку, у лісах мохоподібні часто пригнічують сходи судинних рослин, корені яких не можуть відразу пробити товстий моховий покрив і, перш ніж досягнути ґрунту, піддаються несприятливому впливу чергування сухості й вологості мохової дернини. Деякі лісові мохи виділяють речовини, що гальмують проростання насіння деревних рослин. [9]
Тваринний світ біоценозу також робить на мохоподібні прямий або непрямий вплив. Хребетні тварини вживають їх у їжу рідко (наприклад, лемінги в тундрі харчуються поряд із квітковими рослинами і деякими мохами). Але багатьом безхребетним тваринам мохоподібні є не тільки їжею: утворені ними дернини і килими - середовище існування цих тварин. При цьому деякі з безхребетних тварин, очевидно, можуть брати участь у заплідненні мохоподібних, переносячи крапельки слизу зі сперматозоїдами з однієї рослини на іншу. Наземні хребетні впливають на моховий покрив і непрямо: позитивно або негативно. Кал і трупи деяких тварин, як уже відзначалося, можуть слугувати субстратом для певних видів мохів. Тварини можуть також сприяти поширенню мохоподібних, переносячи їхні спори або прилиплі до ніг виводкові бруньки, листочки і т.п. у нові місця. У деяких мохів навіть є особливі пристосування для залучення тварин, що беруть участь у поширенні спор. Так, у сплахнових, що часто поселяються на екскрементах жуйних, гіпофіза при дозріванні коробочки нерідко сильно збільшується в розмірах і зафарбовується в яскравий колір, а спорова маса має сильний неприємний запах. Мухи, що відкладають свої яйця в гній, приваблюються гіпофізою і (або) запахом, сідають на коробочку. Ковзаючи по ній, вони доторкаються до липкої маси спор, що виступає з коробочки. Потім у пошуках субстрату для відкладання яєць мухи перелітають в інші місця і можуть перенести прилиплі до них спори сплахнових на нові купки екскрементів.
Мохоподібні випробують на собі також величезний прямий і побічний вплив господарської діяльності людини (вирубка і посадка лісів, створення пасовищ і сінокосіння, затоплення й осушення територій, механічна і хімічна обробка ґрунту, будівельні роботи і т.д.). Людина впливає на моховий покрив просто ущільнюючи ґрунт при ходьбі. Навіть слабке порушення підстилки в хвойних лісах веде до відмирання багатьох видів мохів, а подальше витоптування може призвести і до зміни одного біоценозу іншим.
Розглянутий зв'язок мохоподібних з різними факторами зовнішнього середовища пояснює, чому окремі мохоподібні або їхні угруповання можна зустріти в різних кліматичних поясах (від екваторіального до арктичного) і на різних висотах над рівнем моря (від глибоких низин до альпійського пояса). Найбільше число видів мохоподібних знаходяться у дощових тропічних лісів, де майже усі вони є епіфітами. При просуванні в більш високі пояси число видів мохоподібних зменшується, але роль окремих видів у рослинному покриві часто значно зростає в силу їх більшої, в порівнянні із судинними рослинами, пристосованості до умов існування в арктичному поясі.
Різноманітною є ценотична приуроченість виявлених мохоподібних. Так, частина з них відмічена нами лише у соснових фітоценозах. Це Dicranum polysetum, Dicranum scoparium, Polytrichym juniperinum, Hypnum cupressiforme, Brachythecium oldipodium, Pleurozium Schreberi. У всіх обстежених лісових масивах відмічені Ceratodon purpureus, Funaria hygrometrica та Pohlia nutans. Решта видів Atrichum undulatum, Ceratodon purpureus, Mnium cuspidatum, Platygyrium repens, Pylaisia polyantha, Hypnum pallescens, Eurhynchium hians відмічені лише у листяному лісі. Причому вони входять до складу епігейних, епіксильних та епіфіт них бріоугрупувань. Так види - епіфіти Pylaisia polyantha, Hypnum pallescens, Platygyrium repens утворюють епіфітні обростання на стовбурах дерев. В їх прикореневій частині такі угруповання сформовані факультативним епіфітом Mnium cuspidatum. Епігейні бріоугруповання в листяному лісі утворені Atrichum undulatum, Ceratodon purpureus, Mnium cuspidatum.
Отже, мохоподібні маючи своєрідну будову, різну вимогливість до умов середовища, належить до різних екогрупи по відношенню до світла, вологи та ін.
В процесі онтогенезу і під сукупною дією екофакторів у рослин формується своєрідні життєві форми. Аналіз життєвих форм мохоподібних показав, що найбільш багато чисельною є група мохів, які формують пухку дернинку.
Розділ 6. Господарське значення мохоподібних і їхня роль у природі
Малопомітні і непривабливі, на перший погляд, мохоподібні відіграють велику і важливу роль у житті природи. Уловлюючи енергію Сонця, виділяючи кисень, беручи участь у кругообізі речовин й енергії на Землі, як і інші рослини, являють собою незамінний компонент біосфери Землі, невід'ємною частиною якої є і людина.
Здатні переносити різкі коливання температури, надлишкове зволоження або жорстокі посухи, пристосовані до життя на бідних субстратах, вони утворюють угруповання в таких місцях, де вищі судинні рослини пригноблені або зовсім не можуть існувати. Мохоподібні звичайно входять до складу первинних рослинних угруповань на поверхні скель і каменів, вони часто являються піонерами заростання знижень, заповнених водою, і незаселених ґрунтів. Поступово відмираючи, піонерні види мохоподібних готують субстрат для поселення інших видів бріофітів або судинних рослин.
Іноді роль мохоподібних в угрупованнях підвищується в зв'язку з різкою зміною умов місцеперебування, викликаною впливом тварин, людини і факторів, обумовлених життєдіяльністю самої сукупності рослин. Наприклад, знищення лісу - цього могутнього насоса, що викачує вологу з ґрунту, у безстічних місцях або в місцях зі слабким стоком і близьким до поверхні рівнем ґрунтових вод призводить до появи в ґрунті надлишкової вологи. Починається процес заболочування. На ґрунті, покритому до вирубки, наприклад, чорницею, лісовими травами і деякими лісовими мохами, з'являється суцільний покрив з моху політрихума звичайного (Polytrіchum commune). Потім з'являються сфагни, роль яких у рослинному покриві поступово зростає, і нарешті утвориться мохове болото. При поновленні лісу порушений баланс вологи в ґрунті відновлюється, сфагнові і політрихові мохи поступаються місцем іншим, характерним для лісу мохам і квітковим рослинам. Заболочування лісів може мати і зворотній характер. Звичайно такий процес спостерігається по периферії великих сфагнових боліт, що збільшуються і поширюються в зв'язку із загально-кліматичними змінами.
На півночі тайгової зони, там, де кількість опадів, що випадають, досить велика, нерідко відбувається заболочування луків. Накопичена при відмиранні трав органічна речовина в умовах вологого і прохолодного клімату не встигає розкладатися і перетворюється в торф'яний шар, на якому поселяються мохи, згодом утворюють суцільний килим. Якщо людина не втрутиться в життя луків, не поліпшить аерацію і живильний режим ґрунту, вони можуть перетворитись в болото.
Широко відома роль мохоподібних у складі рослинного покриву сфагнових боліт. Але роль боліт не зводиться лише до того, що на них добувають торф, який використовується в якості палива, сировини для хімічної промисловості, будівельного матеріалу й у сільському господарстві.
Болота - це і місце існування багатьох, у тому числі промислових тварин, і невикористані поки належною мірою ягідні угіддя. Особливо велика роль їх (верхових сфагнових боліт) як резервуарів прісної води і регуляторів гідрологічного режиму території. Всмоктуючи в себе як губка літні і зимові опади, болота поступово віддають їх протягом року ґрунтові і струмочкам, що беруть з них початок - джерелам великих рік. Нерідко осушення навіть невеликих боліт, продиктоване господарською потребою (меліорація ділянки лісу або заплави, одержання торфу для добрива і т.п.) обертається великими втратами не тільки для живої природи, але і для народного господарства - обмілінням і замуленням річок, зниженням рівня ґрунтових вод на вже існуючих угіддях.
Велика роль мохоподібних у збереженні природної рівноваги в районах поширення вічної мерзлоти - у тундрі, лісотундрі, тайзі, особливо на ділянках, вкладених пухкими ґрунтами. У таких місцях суцільний мохово-лишайниковий покрив і невеликий торф'янистий шар захищають ґрунти від нагрівання сонячними променями. Порушення цілісності мохового покриву (при пожежах, проїзді транспорту і вирубці лісу в теплий час року, будівництві і т.д.) веде до збільшення глибини, посилення мерзлотних процесів у ґрунтах. Часто при цьому відбуваються катастрофічні, а головне, необоротні зміни ландшафту. На місці стійких форм рельєфу утворяться яри, термокарстові озерно-болотні балки, виникають зсуви. Скорочуються площі лісів, цінних пасовищних і мисливських угідь, руйнуються спорудження. Усе це необхідно враховувати при господарському освоєнні території Крайньої Півночі і Сибіру. [4]
Безпосередньо використовуються людиною лише деякі мохоподібні. Сфагнуми, що володіють антибіотичними властивостями, великою вологоємністю і гарними теплоізоляційними якостями, вживалися в медицині для перев'язок і замість пелюшок при догляді за дітьми. Використовувався djyb в якості теплоізоляційного матеріалу в будівництві. Річчія плаваюча і деякі мохи вирощуються в акваріумах. Маршанція мінлива (Marchantіa polymorpha) і фунария djkjujvshyf (Funarіa hygrometrіca) - улюблені об'єкти експериментальної ботаніки в дослідах по вивченню впливу різних факторів зовнішнього середовища на рослини. Мохоподібні використовуються також як індикатори забруднення атмосфери, глибини відтавання ґрунту і порушення умов життя в біоценозах. У таких випадках, обстежуючи флору мохоподібних якого-небудь району, звертають увагу на наявність або відсутність у ній видів, що характеризуються уже відомим ступенем чутливості до домішок в повітрі сірчистого газу, до заболочування території. Отже роль мохоподібних в природі та житті людини є важливою та багатогранною.
Висновки
1. У результаті наших досліджень було зібрано 19 видів мохоподібних, які належать до 7 родин, 11 родів та 15 видів. Найбагатшою за видовим складом є родина Hypnaceae - 4 види (Platygyrium repens, Pylaisia polyantha, Hypnum pallescens, cupressiforme), а також родина Dicranaceae - 3 види (Dicranum polysetum, scoparium, eratodon purpureus).
Двома видами представлені родини: Polytrichaceae (Atrichum undulatum, Polytrichum juniperinum); Brachytheciaceae (Brachythecium oldipodium, Eurhynchium hians). Родини Bryaceae (Pohlia nutans), Mniaceae (Mnium cuspidatum), Hypnaceae (Platygyrium repens), Funariaceae (Funaria h grometrica) представленні одним видом. В цілому спостерігається перевага родин, види яких приурочені до лісових фітоценозів.
2. За відношенням рослин до вологи виділяють такі екогрупи: гідато-, гідро-, гігро-, мезо- і ксерофіти, а також проміжні групи між ними. Мохоподібні виявлені нами належать до двох груп: мезоксерофіт (60% виявлених мохів) і мезофіти (40%). У залежності від типу субстрату на якому поселяються бріофіти, останні поділяються на епіфіти, епігеї, епік сили, епіліти. Серед виявлених нами мохоподібних є епігеї (11 видів), епіфіти (7 видів), епіксили (8 видів).
3. Велику роль у житті мохоподібних, як і всіх інших рослин відіграє світло. У залежності від його потреб останні поділяються на такі екогрупи: світлолюби, тіневитривалі, тінелюби. Серед зібраних нами видів до світлолюбів належить 4 вида (%), до тіневитривалих 10 видів (%), а до тінелюбів 1 вид (%).
4. В процесі життєдіяльності рослини формують своєрідні життєві форми. У мохів вони характерні не для однієї особи, а для цілої дернинки. Виявлені нами види мають такі життєві форми: пухка дернина (7 видів), подушковидна дернина (1 вид), плоский килим (4 види), плетево (3 види).
5. При дослідженні даної території ми враховували і частоту трапляння певного виду моху. Досить часто ми зустрічали такі види як Дикран віничний, цератодон пурпурний, мній загострений, евринхій зіяючий. Ці види відносяться до різних екогруп за відношенням до умов існування. Зокрема такі види як мній загострений та еврінхій зіяючий були виявлені нами у складі гнізда.
Список використаної літератури
1. Абрамов І.І., Савіч-Любицька Л.І. Основы палеонтологии., М., 1963. - ст.23 - 35;
2. Бардунов Л.В. Древнейшие на суше. - Новосибирск: Наука, 1984. - ст.14-25;
3. Бачурина Г.Ф. Партика Л.Я. Печеночники и мхи Украины и смежных территорий // Киев: Наукова думка, 1979;
4. Большая энциклопедия Кирилла и Мефодия. - М., 1998;
5. Гапон С.В. Байрак О.М. Строгаль Т.Г. // Методичні рекомендації по вивченню мохів та лишайників у природі // Полтава, 1990, ст.7-12;
6. Гапон С.В. Еколого-ценотичні особливості мохоподібних Лівобережного Лісостепу України // Актуальні питання ботаніки та екології // Тези - доповідей конференції молодих учених/ - Полтава, 1992;
7. Гапон С.В. мохоподібні епіфіт них обростань // Укр. бот. Журн. - 1992., ст.13-15;
8. Гарибова Л.В. Дундин Ю.К. Коптяева Т.Ф. Филин В.Р. - Водоросли, лишайники и мохообразные СССР. - М.,: Мысль, 1978;
9. Еленевский А.Г. Соловьева М.П. Тихомиров В.Н. Ботаника систематика высших, или наземных, растений. - М,: Академия, 2004, ст.44-65;
10. Лазаренко А.С. Определитель лиственных мхов Украины // К.: изд. - во А.Н. УССР. - 1955. - ст.15-40;
11. Мальовнича Полтавщина: путівник по заповідниках і пам’ятках природи. -Х.: Прапор, 1982;
12. Маца К.О., Чичкало Б.В., Коваленко Г.М. - Полтава: обл. управл. по пресі, 1998;
13. Мейер К.И. Размножение растений. - М., 1937;
14. Мельничук В.М. Определитель лиственных мхов средней полосы и Юга Европейской части СССР. - К.: Наук. думка, 1970. - ст.10-32;
15. Полтавська область: природа, населення, господарство: Географічний та історико-екологічний нарис.
16. Полтавщина; Енциклопедичний довідник / за ред. А.В. Кудрицького. - К.: УЕ, 1992.;
17. Тахтаджян А.Л. Высшие растения., Т.1, М. - Л., 1956;
18. Чернова Н.М., Билова О.М. Екологія. - К.: Вища школа, 1986.