
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Рефераты по косметологии
Рефераты по криминалистике
Рефераты по криминологии
Рефераты по науке и технике
Рефераты по кулинарии
Рефераты по культурологии
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Рефераты по косметологии
Рефераты по криминалистике
Рефераты по криминологии
Рефераты по науке и технике
Рефераты по кулинарии
Рефераты по культурологии
Курсовая работа: Живлення рослин вуглецем
Курсовая работа: Живлення рослин вуглецем
ЗМІСТ
Вступ. 2
Розділ 1. Значення фотосинтезу. 3
1.1 Суть та значення фотосинтезу. 3
1.2 Загальне рівняння фотосинтезу та походження кисню.. 7
Розділ 2. Листок як орган фотосинтезу. 13
Розділ 3. Фотосинтетичні пігменти листка. 19
3.1 Хлорофіли. 19
3.2 Каротиноїди. 22
3.3 Фікобіліни. 24
3.4 Оптичні властивості фотосинтетичних пігментів. 27
3.5 Біосинтез пігментів фотосинтетичного апарату рослин. 28
Розділ 4. Енергетика фотосинтезу. 33
4.1 Первинні процеси фотосинтезу. 33
4.2 Фотосинтетична одиниця, реакційний центр, фотосистема. 36
4.3 Z-cxeмa фотосинтезу. 40
Висновки. 43
Список літератури. 45
Вступ
Нині людство все більше розуміє ту очевидну істину, яку вперше обґрунтували видатні вчені К. А. Тімірязєв і В. І. Вернадський, що екологічне благополуччя біосфери, а отже й існування самої людини та цивілізації в цілому, визначається станом рослинного покриву планети. Значення фотосинтезу в біосферних процесах Землі настільки велике й різноманітне, а його природа настільки унікальна, що проблема фотосинтезу правомірно вважається однією з найважливіших проблем не лише науки, а й практики. Більш як 3,5 млрд. років тому відбувалася подія, якій судилося стати могутньою рушійною силою еволюції органічної матерії — виник фотосинтез. Еволюційний процес випробував різні варіанти енергозабезпечення життя. Перші прокаріоти типу бактерій були ферментуючими гетеротрофами, які добували їжу шляхом розщеплення органічних речовин абіотичного походження. Життя набуло якісно нове, практично невичерпне джерело енергії — Сонце.
Прокаріоти — це ціанобактерії, що дали початок розвитку рослинного царства, знайшли ключ до фотоавтотрофії. Центральне місце в цьому процесі зайняв зелений пігмент — хлорофіл. Один з основоположників фізіології рослин К. А. Тімірязєв, підкреслюючи космічне значення зеленої рослини, образно писав, що зелений листок, а вірніше хлорофіл, є фокусом у світовому просторі, в який, з одного боку, йде енергія Сонця, а з другого — беруть початок усі прояви життя на Землі.
Мета роботи охарактеризувати протікання процесів живлення рослин вуглецем.
Завдання роботи:
1) розглянути значення та суть процесу фотосинтезу;
2) дати характеристику листку як органу фотосинтезу;
3) дати характеристику основних фото синтезуючих пігментів листка;
4) розглянути енергетичні процеси фотосинтезу.
Розділ 1. Значення фотосинтезу
1.1 Суть та значення фотосинтезу
Фотосинтез — єдиний процес у біосфері, який веде до збільшення вільної енергії біосфери за рахунок зовнішнього джерела — Сонця і забезпечує існування як рослин, так і всіх гетеротрофних організмів, у тому числі й людини.
Зараз важко, а то і зовсім неможливо знайти будь-які природні явища, які не були б пов'язані з фотосинтезом. Таким чином, основним джерелом енергії для фотосинтезу та біологічної продуктивності слугує енергія Сонця. Рослини перехоплюють цю енергію, але використовують для фотосинтезу лише 2...5 %, решта витрачається на нагрівання рослин і довкілля, тому сонячна енергія визначає також температуру, за якої відбуваються фізіологічні процеси. Крім фотосинтезу, сонячна радіація (енергія, яка переноситься в просторі у формі електромагнітних хвиль або квантів) впливає також на фотоморфогенетичні та фототропні реакції.
Фотосинтетично активна радіація (ФАР) — це радіація в межах 400...700 нм, вона вимірюється в моль • м-2* с, або Вт * м-2, тоді як енергія квантів різних довжин хвиль — в кДж або в еВ (табл. 1).
Таблиця 1. Енергія квантів різних довжин хвиль
| Довжини хвиль (нм) | Колір світла | Вміст енергії кДж (моль фотонів) | ДЕ(еВ) |
| 700 | Червоний | 170 | 1,76 |
| 650 | Оранжевий | 183 | 1,90 |
| 600 | Жовтий | 199 | 2,06 |
| 500 | Зелено-синій | 238 | 2,47 |
| 440 | Блакитний | 271 | 2,80 |
| 400 | Фіолетовий | 298 | 3,09 |
Кванти світла (фотони) в даній області спектра майже однаково ефективні в фотосинтетичних реакціях. ФАР становить близько 50 % загальної (пряма + розсіяна) сонячної радіації й дещо змінюється залежно від часу доби чи пори року. Радіація, що падає на рослинний покрив, може поглинатися, передаватися чи відбиватися. В спектральній області ФАР листок поглинає до 80 % падаючої радіації, тоді як у короткохвильовій та інфрачервоній областях більша частина її розсіюється. Значення цього явища полягає в тому, щоб зменшити теплове навантаження від хвиль тих довжин, які не використовуються у фотосинтезі. Однак у деякій інфрачервоній області листки добре поглинають радіацію. Оскільки листки одночасно є і добрими джерелами випромінення, вони здатні досить ефективно розсіювати надлишок тепла в довгохвильовій частині спектра. За період еволюції протягом мільярдів років природа створила процеси та живі системи, які ідеально використовують потоки енергії сонячної радіації.
Маса рослинного покриву Землі складає більш як 1800 млрд. т сухої речовини, що енергетично еквівалентно 30 • 1021 Дж. Ця цифра відповідає відомим запасам енергії корисних копалин. Ліси становлять близько 68 % біомаси суші, трав'янисті екосистеми — 16, агрофітоценози — лише 8 %. В цілому на Землі з участю фотосинтезу щороку створюється 173 млрд. т сухої речовини.
Енергетичний еквівалент щорічної біопродуктивності на Землі приблизно в 10 разів більший за річну енергетичну потребу всього населення земної кулі.
Поступово, всього лише сотні мільйонів років тому, кількість первинної продукції живих організмів на Землі зрівноважилася з її витратами в процесах дихання, бродіння, окиснення. На планеті стабілізувався добре врівноважений кругообіг вуглецю та інших органогенів, насамперед азоту, фосфору, калію, що стало одним із найважливіших факторів формування оптимального для життя екологічного середовища.
Кругообіг кисню, вуглекислого газу, азоту та інших елементів, які беруть участь у процесах життєдіяльності рослин, створив і підтримує газовий склад сучасної атмосфери, потрібний для життя на Землі.
Однак науково необґрунтована інтенсифікація діяльності людства, поява нових екологічно небезпечних джерел енергії, наприклад атомного розпаду, призвели до сучасної екологічної кризи. Вміст СО2 в атмосфері почав збільшуватися, що може різко вплинути на тепловий режим планети. СО2 та водяна пара поглинають інфрачервоні промені, що відбиваються від Землі в космос і створюють так званий парниковий ефект. За даними агентства з охорони природи США, це загрожує підвищенням температури на планеті в найближче століття на 3...9 °С із наступними катастрофічними наслідками. Фотосинтез перешкоджає нагромадженню СО2, захищає планету від перегріву. Крім того, постійний вміст кисню (21 %) в атмосфері забезпечує існування озонового екрану на висоті 25 км. Озон — О3 — створюється в результаті фотодисоціації молекул О2 під дією сонячної радіації. Він захищає все живе від згубної дії ультрафіолетових променів (240...290 нм). Забруднення атмосфери, знищення лісів призводить до зниження вмісту кисню, виникнення озонових дірок. Звідси прямий зв'язок фотосинтезу з охороною навколишнього середовища не в регіональних, а в глобальних масштабах.
Якщо раніше фотосинтез асоціювався з біологією, а правильніше, з фізіологією рослин, то останнім часом ним серйозно зацікавилися технологи. Виявилося, що рослинний організм, здатний перетворювати світлову енергію на електричний струм, вилучає з води водень — цінне, екологічно безпечне, але поки що дороге паливо, фіксує вільний азот повітря та багато іншого, що може бути використане в фітобіотехнології.
Вивчення механізму фотосинтезу дасть змогу здійснювати цей процес у промислових масштабах поза рослиною, що має вирішальне значення в енергетиці майбутнього.
Не менш важливе значення фотосинтезу як основи одержання продовольства, кормів, технічної сировини. Глобально чиста продуктивність фотосинтезу за розрахунками становить 78 • 109 т вуглецю за рік, з яких 7 % — безпосередньо або через тваринні організми використовуються як продукти харчування, паливо, сировина. Використання корисних копалин як палива експоненційно зростало на 4,3 % за рік протягом останнього сторіччя й на початок XXI ст. приблизно зрівнялося з наростанням біомаси.
Незважаючи на високу ефективність початкових стадій фотосинтезу (95 %), в урожай переходить менш як 1...2 % сонячної енергії, втрати якої зумовлені неповним поглинанням світлових променів, лімітуванням процесів на рівні біохімії та фізіології. Рослинність всіх континентів фіксує у вигляді хімічної енергії приблизно 1...2 % ФАР.
Згідно з теоретичними розрахунками, максимальна енергетична ефективність може бути до 28 % ФАР. При коефіцієнті корисної дії 3...5 % ФАР можна одержати до 60 т/га органічної маси. В зв'язку з цим особливо актуальними стають питання розробки теоретичних основ управління фотосинтезом, дослідженням його як єдиного процесу запасання сонячної енергії, закономірностей його регулювання та адаптації до зовнішніх умов. Визначальна залежність людства від фотосинтезу особливо проявляється в наш час, коли невідповідність між виробництвом сільськогосподарської продукції та чисельністю народонаселення стає все більш загрозливою.
Подальше інтенсивне дослідження законів еволюції біосфери у зв'язку з фотосинтетичним процесом, їх прогнозування, приведення в агрокліматичну відповідність різних регіонів планети вселяє впевненість, що за оптимальних умов агрофітоценози зможуть забезпечити їжею більш як 10... 12 млрд. чоловік — найнижчий імовірний рівень населення нашої планети. Однак для цього слід перейти до ери стабільного рівномірно розподіленого споживання продуктів харчування на помірному рівні, який дав би змогу зберегти буферну здатність глобального циклу вуглецю.
І нарешті, слід підкреслити світоглядне значення фотосинтезу, бо вивчення цього процесу має привести нас до розкриття кардинальної проблеми — з'ясування того, як виникло життя на Землі. Адже в процесі фотосинтезу щомиті неорганічна речовина СО2 та Н2О перетворюються на органічну, а, як відомо, саме в цьому і полягає суть переходу від передбіологічної еволюції до біологічної. Отже, фотосинтез як явище і наукова проблема — екстраординарний.
З нагромадженням знань про особливості даного процесу змінювалась і його назва. Тепер вважають недостатнім класичне визначення, згідно з яким фотосинтез — це утворення зеленими рослинами органічних сполук із СО2 і Н2О за допомогою квантів сонячного світла. В наш час фотосинтез визначають як фототрофну функцію бактерій, найпростіших, водоростей та вищих рослин.
Фототрофна функція — це сукупність процесів поглинання, перетворення та використання в багатьох ендорганічних реакціях світлових квантів, у ході яких відбувається первинне становлення пластичних та енергетичних ресурсів життя на нашій планеті.
1.2 Загальне рівняння фотосинтезу та походження кисню
Датою відкриття процесу фотосинтезу вважають 1771 рік. Англійський вчений Д. Прістлі звернув увагу на зміну складу повітря внаслідок життєдіяльності тварин. У присутності зелених рослин повітря знову ставало придатним як для дихання, так і для горіння. В подальших роботах учених (Й. Інгенхауз, Ж. Сенеб'є, Н. Соссюр, Ж. Бусенго) було встановлено, що зелені рослини із повітря поглинають вуглекислий газ, із якого за участі води на світлі утворюється органічна речовина. Саме цей процес у 1877 р. німецький вчений В. Пфеффер назвав фотосинтезом. Величезне значення для розкриття суті фотосинтезу мав закон збереження енергії, сформульований Р. Майєром (1845). Згідно з цим законом, енергія, яку використовують рослини, — це енергія Сонця, яку в процесі фотосинтезу рослини перетворюють на хімічну енергію.
Загальне рівняння фотосинтезу не досить точно передає суть цього процесу, адже, згідно з цим рівнянням, можна припустити, що частина виділеного О2 походить від СО2, тоді як насправді весь кисень утворюється з води в ході фотолізу:
![]()
Тому доцільніше розділити його на складові частини:
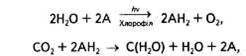
де А — акцептор електронів і протонів.
Згідно з цим рівнянням, в основі суті фотосинтезу лежить окисно-відновна реакція. За участі хлорофілу та енергії сонячних квантів вода фотоокиснюється, в результаті чого виділяються кисень та водень, останній і відновлює СО2 до рівня вуглеводів. Ці реакції відбуваються відповідно в світлову та темпову стадії фотосинтезу.
Впродовж сторіччя після відкриття Д. Прістлі (1771) фотосинтезу вчені з'ясовували його суть, встановили учасників і кінцеві продукти, що одержуються в результаті цього процесу.
Довгий час вважали загальноприйнятим, що вуглеводи утворюються з вуглецю та водню, а кисень виділяється з вуглекислоти. Ця гіпотеза, як виявилося пізніше, була абсолютно неправильною. Вперше в 1893 р. О. М. Бах передбачив, що асиміляція СО2 в процесі фотосинтезу нічого спільного з відщепленням кисню від СО2 не має, а пов'язана з окисно-відновним процесом, який відбувається з участю водню та гідроксилу води, причому кисень виділяється з води.
Теорію походження кисню з вуглекислого газу спростував Корнеліус Ван Ніль, який в Станфордському університеті (США) вивчав метаболізм різних фотосинтезуючих бактерій. Він з'ясував, що для фотосинтезу пурпурних сірчаних бактерій необхідний сірководень, а в результаті цього процесу всередині бактеріальних клітин нагромаджується сірка:
![]()
Після серії дослідів у 1937—1941 pp. учений зробив сміливе узагальнення: первинна фотохімічна реакція фотосинтезу полягає саме в фотодисоціації води, а не в розкладанні СО2, й запропонував сумарне рівняння фотосинтезу:
![]()
В цьому рівнянні Н2А може бути водою або іншим відновником. Отже, бактерії, здатні до фотосинтетичної асиміляції СО2, за винятком ціанел, використовують H2S, H2, СН3, а тому в процесі фотосинтезу не виділяють кисень. Такий тип фототрофного живлення дістав назву фоторедукції. Для вищих рослин і водоростей Н2А — це вода. Інакше кажучи, Ван Ніль припустив, що саме вона, а не вуглекислий газ розкладається в процесі фотосинтезу. Отже, рівняння фотосинтезу тепер матиме такий вигляд:
![]()
або поетапно:
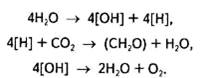
Уявлення Ван Ніля про походження кисню знайшло підтвердження в експериментах із міченим 18О2, які незалежно в 1941 р. провели С. Рубен (США) та російські вчені О. П. Виноградов і Р. В. Тейс. За допомогою мас-спектрометричного аналізу було показано, що відношення 16О2 до 18О2 в кисні, який утворюється в процесі фотосинтезу, відповідає співвідношенню їх саме у воді, а не в СО2:
![]()
Коли ж ввести ізотоп О2 у вуглекислий газ, то рівняння набуває іншого вигляду:
![]()
Це свідчить, що новоутворена молекула води відрізняється від тих двох молекул, що вступили в реакцію (рис. 1).
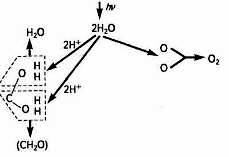
Рис. 1. Фотоліз води і виділення О2
Отже, порівняльний аналіз ізотопного складу води, вуглекислого газу та О2, що виділяється під час фотосинтетичного процесу рослиною, підтвердив спорідненість кисню фотосинтезу з водою.
Також Р. Хілл у 1939 р. продемонстрував, що ізольовані хлоропласти здатні під час освітлення розкладати воду з виділенням О2 за наявності відповідних акцепторів електронів, але без участі вуглекислого газу:
![]()
Відсутність реакції, в якій були б використані електрони, компенсується, як видно з реакції, введенням у систему акцептора електронів (А), або «реагента Хілла». Це можуть бути солі заліза (калій феріоксалат, калій фериціанід), бензохінон або природний акцептор електронів НАДФ+.
Отже, для водоростей, вищих рослин, у яких вода слугує донором електронів, підсумкове рівняння фотосинтезу матиме такий вигляд:
![]()
При цьому також запасається вільна енергія, величина якої для цього процесу приблизно 470 кДж/моль.
Під час фотосинтезу з простих неорганічних сполук (води та вуглекислого газу) синтезуються різноманітні органічні сполуки. В результаті відбувається перебудова хімічних зв'язків: замість С—О та Н—О виникають С—С та С—Н зв'язки, в яких електрони займають більш високий енергетичний рівень.
Дослідження свідчать також, що майже весь кисень атмосфери має фотосинтетичне походження. Отже, процеси дихання і горіння стали можливі лише після того, як виник фотосинтез. Все це й свідчить про космічне значення зеленої рослини. Підсумкова реакція фотосинтезу може мати вигляд окисно-відновних реакцій, одна з яких фотодисоціює воду, а інша — відновлює СО2 до вуглеводів, але вже без участі квантів сонячних променів:
![]()
І дійсно, нині відомо, що фотосинтез відбувається в дві стадії — світлову та темнову.
Розділ 2. Листок як орган фотосинтезу
Особливості морфології, анатомії та фізіології листка повною мірою забезпечують здійснення його основної функції — фотосинтезу (рис. 2).
Загальна листкова поверхня рослин помірної зони, розміщених на 1 га орної площі, майже в п'ять разів перевищує земельну ділянку, а в південних районах з нормальним зволоженням — у 10... 15 разів. Різноманітна їхня форма, розташування на стеблі, фото-тропічна реакція забезпечують виконання найважливішої функції — поглинання світлових променів. Як і всі фізичні тіла, листок відбиває, поглинає та пропускає промені, що падають на нього (рис. 3).
Поверхня листка вкрита епідермісом. Клітини нижнього та верхнього епідермісу, за винятком тих, що створюють продиховий апарат, позбавлені хлоропластів. Вони лише пропускають світлові промені, отже, безпосередньої участі в фотосинтезі не беруть.
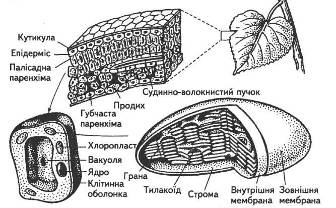
Рис. 2. Листок як орган фотосинтезу
Характерною особливістю покривної тканини листка є наявність продихів, площа яких не перевищує 1 % його загальної поверхні. Для того щоб процес фотосинтезу протікав нормально, в клітини до зелених пластид має надходити вуглекислий газ. Кількість його в атмосферному повітрі становить близько 0,03 %. Впродовж дня рослина засвоює таку кількість СО2, яка міститься приблизно в 30...60-метровому шарі повітря. Для утворення 1 г вуглеводів необхідно близько 1,47 г СО2 (така кількість його міститься майже у 2500 л повітря). Це можливо завдяки неперервному турбулентному рухові повітряних мас навкруг листків, який спричинюється нерівномірним нагріванням їх сонячними променями та вітром.
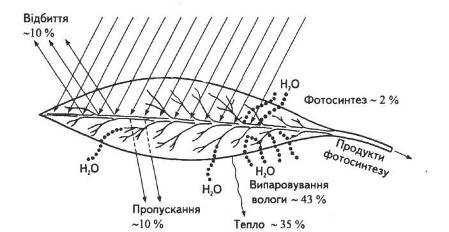
Рис. 3. Енергетичний баланс листка
Кількість води, що витрачається на фотосинтез, становить зовсім незначну частину тієї кількості, яку поглинає та випаровує рослинний організм. Дефіцит води знижує швидкість фотосинтезу, адже закриття продихів припиняє доступ вуглекислого газу до мезофілу листка. Крім того, кисень, що виділяється в процесі фотосинтетичних реакцій фотолізу води, теж потрапляє в навколишнє середовище крізь продихи. Тому закриття продихів припиняє газообмін з атмосферою, хоча при цьому ні фотосинтез, ні дихання не припиняються. Як невід'ємні структури листка, продихи взаємодоповнюють один одного, постачаючи відповідно кисень та вуглекислий газ для відповідних реакцій.
Отже, функціонування продихів регулює газообмін — надходження вуглекислого газу, виділення кисню та випаровування води. Як правило, продихів більше з нижнього боку листка, куди не потрапляють прямі сонячні промені. На верхній частині їх значно менше, що зменшує випаровування води під дією тих самих променів.
Основну ж тканину листка займає мезофіл, який буває двох типів: губчастий та палісадний (рис. 4).
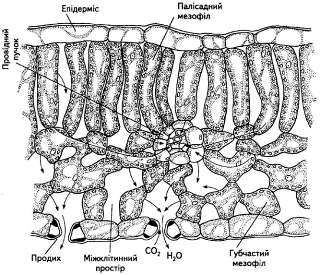
Рис. 4. Анатомічна структура листка
Найбільше хлоропластів мають клітини палісадної або, як її ще називають, стовбчастої паренхіми (кілька десятків на одну клітину). Ці клітини щільно прилягають одна до одної, на відміну від пухко розміщених клітин губчастого мезофілу. Від 15 до 20 % загального об'єму листка становить міжклітинний простір, наявність якого характерна насамперед для губчастої паренхіми. Завдяки міжклітинникам значно зростає внутрішня поверхня листка (вона в 7... 10 разів більша за зовнішню поверхню), створюється відповідне співвідношення газового складу, яке відрізняється від атмосферного, хоча міжклітинний простір і поєднується з повітрям атмосфери через продихи. Саме за рахунок міжклітинного простору СО2 постійно надходить з атмосфери, бо губчаста тканина мезофілу розміщується поблизу нижнього епідермісу, де розташовані численні продихи.
Мезофіл листка пронизаний густою сіткою судинно-волокнистих пучків, до складу яких входить ксилема, яка постачає воду та мінеральні речовини, і флоема, яка забезпечує відтік продуктів фотосинтезу.
Характерно, що прямого контакту між клітинами флоеми та кожною клітиною паренхіми немає. Це свідчить про те, що внутрішнє транспортування всього необхідного для фотосинтезу, як і відтік його продуктів на короткі відстані, здійснюється за участі симпласту й апопласту.
Для здійснення фотосинтезу в оптимальному режимі, листок має одержувати достатню кількість світлової енергії, води та вуглекислого газу.
Фотосинтез здійснюється під час безперервного надходження значної кількості світлової енергії в певні ділянки клітин. Тому в процесі еволюції рослин в клітині сформувалися спеціалізовані структури — хлоропласти (рис. 5).
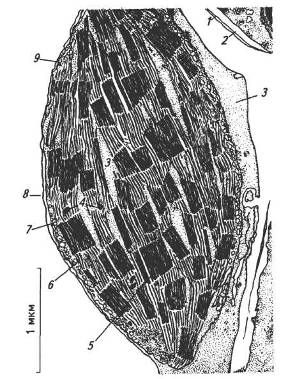
Рис. 5. Будова хлоропласта:
1 — зовнішня оболонка; 2— внутрішня оболонка; 3 — строма; 4 — краплини ліпідів; 5— грана; 6 — тилакоїди строми; 7 — крохмальні зерна; 8 — оболонка хлоропласта; 9 — рибосоми
Довжина хлоропластів коливається від 4 до 10 мкм. Кількість їх у клітині може коливатися від одного до 100 і більше. Загальна сумарна поверхня хлоропластів перевищує площу листків у десятки, навіть у сотні разів.
Хлоропласти — це напівавтономна, саморегулювальна система рослинної клітини, яка для розвитку та функціонування використовує генетичну інформацію як свого власного, так і ядерного, а можливо і мітохондріального, геномів.
У 1960 р. японський біохімік Т. Івамура виявив в одноклітинній водорості хлорелі два типи ДНК — ядерну та хлоропластну. Стало очевидним, що генетичний матеріал евкаріот локалізований не лише в ядрі, яке слугує основною базою збереження спадкової інформації, а й розсіяний по клітині в її субклітинних структурах.
Хлоропластна ДНК у вигляді подвійної спіралі має лінійну, кільце- та петлеподібну форми. Загальна кількість ДНК у хлоропласті залежно від виду коливається в межах 20... 10 • 10-15 г, а молекулярна маса — в межах 84... 104 мегадальтон. Контурна довжина кільцеподібних молекул хлоропластної ДНК у вищих рослин досягає 43...55 мм. В такій ДНК може бути близько 2 • 107 пар нуклеотидів, що відповідає наявності кількох тисяч генів.
Однак генетична інформація пластид досить обмежена. Тому багато компонентів пластид синтезуються під контролем як хлоропластної, так і ядерної ДНК. У хлоропластах є своя власна рибосомальна рРНК, а можливо, і повний набір своїх транспортних тРНК, які беруть участь в експресії геному організму. В стромі хлоропласта локалізовані дрібні рибосоми з коефіцієнтом седиментації 70S. Маючи власний спадковий апарат, власну систему, що синтезує білок, хлоропласт може контролювати синтез певних поліпептидів.
За хімічним складом основна маса хлоропластів складається (% на суху речовину) з: білків (30...60), ліпідів (20...40), хлорофілів (5...9), каротиноїдів (4...5), ДНК (0,01...0,02), РИК (О,5...3,5), мінеральних речовин (6...10). У хлоропластах зосереджено до 80 % усього заліза рослинної клітини, 65...70 % всього цинку та близько 50 % міді. Хлоропласти містять різноманітні ферменти, які забезпечують функціонування ензиматичних реакцій фотосинтезу. В них містяться всі ферменти, які беруть участь в процесі фотосинтезу (рибулозобісфосфат-карбоксилаза/оксигеназа, окисно-відновні, синтетази, гідролази та інші).
Одним із найважливіших компонентів фотосинтетичного апарату є пігментний комплекс. Прокаріоти не мають спеціалізованих фотосинтетичних органел. їхній пігментний апарат представлений в окремих мембранах, які дифузно розсіяні в клітині. Всі інші представники евкаріотів мають спеціалізовані органели — хлоропласти, де й зосереджено весь пігментний комплекс. Виявлено, що фізико-хімічні властивості пігментів найкраще реалізуються саме в хлоропластах.
Розділ 3. Фотосинтетичні пігменти листка
3.1 Хлорофіли
Хлорофіли — це складні ефіри дикарбонової кислоти хлорофіліну, в якої одна карбоксильна група етирифікована залишком метилового спирту, а і інша — залишком спирту фітолу.
Методом хроматографічного аналізу хлорофіли було розділено на хлорофіл а — C55H72O5N4Mg Ta хлорофіл b — C55H70O6N4Mg.
Основу молекули становить порфіринове ядро, що складається з чотирьох пірольних кілець, атоми азоту яких сполучені в центрі магнієм. По периферії пірольні кільця зв'язані між собою метино-вими містками (=СН—). Крім того, є додаткове п'яте циклопента-нове кільце, яке містить кетогрупу (=С=О).
Гідрофільні властивості хлорофілу пов'язані з його азотовмісним порфіриновим ядром — «головкою» молекули. Довгий вуглецевий ланцюг (фітольний «хвіст»), який приєднується до порфіринової частини молекули, надає хлорофілу гідрофобних властивостей. Здатність фітолу розчинятися в жирах має важливе значення для збереження структури хлоропластів. Саме завдяки наявності залишку фітолу хлорофіли добре розчинні в спирті, бензині, ацетоні, але нерозчинні у воді. Протилежні гідрофільні та гідрофобні властивості цих частин молекули мають важливе значення під час локалізації хлорофілу в ламелах хлоропласта між білками та ліпідами. Структурну формулу хлорофілів наведено на рис. 6.
Хлорофіл а наявний у хлоропластах або в аналогічних структурах усіх організмів, здатних до фотосинтезу, за винятком бактерій які містять бактеріохлорофіл а.
Усі вищі рослини та зелені водорості містять також хлорофіл b Різниця між ними полягає в тому, що до хлорофілу ь біля третього атома вуглецю другого пірольного кільця замість метильної групи (-СН3), яка характерна для хлорофілу а, приєднана альдегідна (-СОН)-група. Ці два хлорофіли відрізняються також за забарвленням: хлорофіл а має синьо-зелений відтінок, тоді як хлорофіл b -жовто-зелений. Вміст хлорофілу а в листку приблизно втроє більший, ніж хлорофілу b.
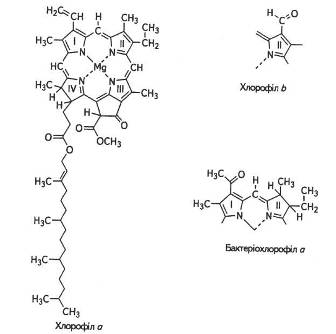
Рис. 6. Структурна формула хлорофілів
Молекула хлорофілу електрично нейтральна, бо подвійний позитивний заряд магнію компенсується двома надлишковими електронами, які розподілені між чотирма атомами азоту пірольних кілець.
Якщо з молекули хлорофілу видалити фітол, одержимо хлорофілід.
У разі заміщення атома Мд2+ воднем утворюється феофітин. У різних представників рослинного світу трапляються й інші види хлорофілів та інших пігментів.
Різні види пігментів розпізнають за допомогою їхніх спектральних характеристик (рис. 7).
Спектри поглинання у різних груп хлорофілів залежать від характеру заміщувальної групи біля пірольних кілець і типу органічного розчинника. Так, бактеріохлорофіл а має спектр поглинання з максимумами 400, 600 і 700 нм. Завжди чітко вирізняються два максимуми поглинання: в області коротких хвиль та відповідно в довгохвильовій. Наприклад, спектри поглинання хлорофілу а — 420 і 662 нм, а хлорофілу b — 455 і 644 нм. В листку спектри поглинання хлорофілу змінюються залежно від його стану, ступеня агрегації йадсорбції на певних білках. Виявлено форми хлорофілів, які поглинають світло з довжиною хвилі 700, 710 і навіть 720 нм.
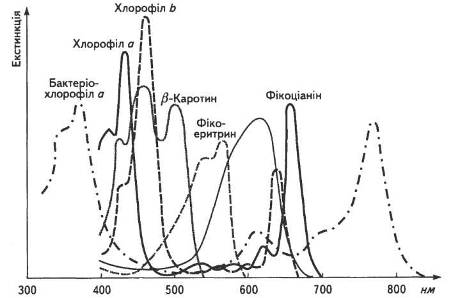
Рис. 7. Спектральні характеристики пігментів
Хлорофіл у живій інтактній клітині здатний до фотоокиснення та фотовідновлення. Здатність до окисно-відновних реакцій пояснюється наявністю в молекулі хлорофілу подвійних зв'язків з рухливими π-електронами та атомів азоту з неподіленими електронами. Азот пірольних кілець може окиснюватися (віддавати електрони) або відновлюватися (приймати електрони).
Важливою властивістю хлорофілів є їхня здатність до взаємодії один з одним, тому в хлоропластах можуть траплятися хлорофіли як у мономерніп, так і в агрегованій формі.
Встановлено, що хлорофіл у мембранах пластид перебуває у вигляді пігмент-ліпопротеїдних комплексів з різним ступенем агрегації. Особливості спектра дії фотосинтезу свідчать про те, що в процесах поглинання світла беруть участь і додаткові пігменти.
3.2 Каротиноїди
Поряд із зеленими пігментами в хлоропластах є й такі, що належать до групи каротиноїдів. Каротиноїди — це найпоширеніші в рослинному світі жиророзчинні жовті, оранжеві та червоні пігменти аліфатичної будови. Вони є обов'язковим компонентом фотосинтетичного апарату. За хімічною природою всі вони — полімери вуглеводню і складають ланцюг із 40 вуглецевих атомів (рис. 4.9), побудований з 8 залишків ізопрену.
Довжина ланцюгів досягає 3 нм і вони часто закінчуються шестичленними циклами. Каротиноїди можуть бути ациклічними (лікопін), моноциклічними або біциклічними. Окиснені форми каротиноїдів називають ксантофілами. Ксантофіли становлять близько 50 % всіх каротиноїдів листка.
У хлоропластах вищих рослин поряд із хлорофілами найчастіше трапляються β-каротин (С40Н56) і ксантофіли — лютеїн (С40Н56О2) і віолаксантин (С40Н56О4). В значних кількостях є також а-каротин і неоксантин.
Каротини, як і ксантофіли, характеризуються гідрофобними властивостями, тому добре розчинні в жирах, що дає їм змогу формувати комплекси в ліпідному прошарку мембрани. Ксантофіли, порівняно з каротинами, мають різноманітнішу структуру, бо до їхнього складу можуть входити різні групи, що містять О2, а саме: гідроксильні, метоксильні, кетогрупи та інші. Між собою ксантофіли відрізняються лише ступенем окиснення.
Як допоміжні пігменти фотосинтетичного апарату каротиноїди забезпечують поглинання квантів двома піками в синьо-фіолетовий та синій областях спектра (420...490 нм) і деякою мірою в зеленій (490...550 нм). Максимуми поглинання залежать від замісників біля вуглецевого скелета, типу розчинника та кількості подвійних зв'язків. В амплітуді від 400 до 550 нм вони, як правило, мають дві-три смуги поглинання.
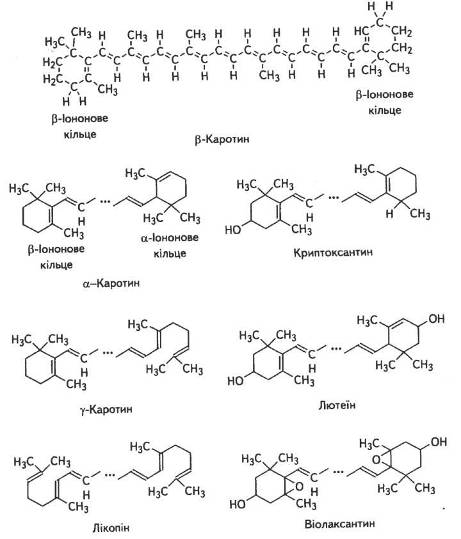
Рис. 8. Структурні формули каротиноїдів
Отже, каротиноїди розширюють спектр дії фотосинтезу, забезпечуючи, поглинання від 10 до 20 % енергії сонячних квантів, причому близько 50 % енергії поглинається в короткохвильовій області — зоні високих енергій. Ці пігменти виконують функцію світлопоглинання, передаючи енергію свого електронно-збудженого стану до хлорофілу а. Зворотний процес передачі неможливий. Слід підкреслити, що каротиноїди, на відміну від хлорофілів, не здатні до флуоресценції.
Такі каротиноїди, як віолаксантин, неоксантин, зеаксантин та інші, поглинаючи світло в короткохвильовій високоенергетичній частині спектра, виконують захисну функцію, як хімічні буфери в реакціях фотосинтезу. Можливий механізм захисту полягає в тому, що каротиноїди здатні реагувати зі збудженою молекулою хлорофілу, забираючи від нього енергію, чим попереджають його фотоокиснення. Енергія фотозбудженої молекули хлорофілу переходить до каротиноїду, хлорофіл набуває нормального енергетичного стану, а енергія виділяється у вигляді тепла. Завдяки цьому каротиноїди оберігають хлорофіл та інші біологічно активні сполуки від фотоокиснення.
Слід згадати, що каротиноїдам належить ще одна специфічна функція в регулюванні фотосинтетичного апарату рослинного організму. Річ у тім, що хлоропласти переміщуються в клітині під впливом синіх променів, які знову ж таки поглинаються каротиноїдами.
Фізіологічна функція каротиноїдів не обмежується їхньою участю в передачі енергії на молекули хлорофілів. Каротиноїди — переносники активного кисню, вони беруть участь в окисно-відновних реакціях завдяки наявності значної кількості подвійних зв'язків. їм належить певна функція у статевому процесі рослин, а саме: вони зумовлюють забарвлення пелюсток квітів, плодів, коренеплодів. Залишається мало з'ясованою їхня функція в кисневому обміні, участь у формуванні фотоперіодичної реакції, в ростових процесах, зокрема під час проростання насіння, в проявах фототаксису та фототропізму.
3.3 Фікобіліни
Такі фотосинтезуючі організми, як синьо-зелені водорості, червоні та деякі інші представники водоростей, окрім хлорофілів і каротиноїдів, мають ще одну групу допоміжних пігментів — фікобіліни. Цю назву вони одержали завдяки своїй подібності до тваринних пігментів — білірубінів.
Білінами називають досить поширені в природі тетрапіроли з відкритим ланцюгом, без металу. В тваринному царстві вони часто трапляються як нефункціональні катаболіти гема. Рослинні біліни (фікобіліни) функціонують як сенсибілізатори і фоторецептори, що забезпечується їхнім ковалентним зв'язком з апобілками. Отже, подібно до хлорофілів, фікобіліни — тетрапіроли, але чотири залишки піролу в них створюють незамкнутий витягнутий або згорнутий ланцюг. Піроли з'єднуються між собою метиленовими і метиновими містками, тоді як хромофорні групи, як правило, ковалентно зв'язані білком.
Хромофорною групою фікоціанінів є фікоціанобілін, фікоерит-ринів — фікоеритробілін (рис. 9).
Відрізняються вони між собою лише взаєморозміщенням пірольних і піроленінових кілець. У фікобілінпротеїнах одна молекула білка може зв'язувати кілька хромофорних груп. Фікоеритробілін у сполученні з білком називають фікоеритрином. Він добре розчинний у воді, поглинає кванти зеленої частини спектра. Фікоеритробіліни (C34H47N4O8) трапляються переважно у червоних водоростей, де й визначають їхнє забарвлення, адже фікоеритрини — білки червоного кольору, які мають максимуми поглинання 500...568 нм.
Фікоціанобіліни (C34H42N4O9) характерні для синьозелених водоростей, причому максимуми поглинання фікоціанінів дещо зсунуті в довгохвильову область — від 585 до 630 нм.
Фікоеритрин і фікоціанін складаються з двох різних білкових субодиниць, які позначають відповідно а (молекулярна маса 19 000) і р (молекулярна маса 21 000) у співвідношенні 1 : 1. Кожна з білкових субодиниць несе ковалентно зв'язаний з нею фікобілін. Як правило, фікобіліни, зв'язані з однією субодиницею, належать до одного типу, тобто до фікоеритробілінів або фіко-ціанобілінів.
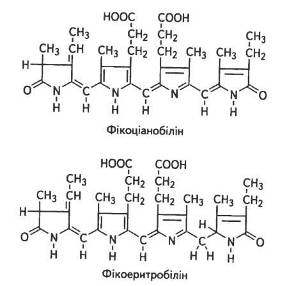
Рис. 9. Хромофорні групи фікоціанінів і фікоеритринів
До фікобілінів належать також алофікоціаніни — пігменти з максимумами поглинання від 585 до 650 нм, хромофорну групу їх поки що не визначено. Цю назву алофікоціаніни одержали завдяки тому, що спочатку їх приймали за одну з форм фікоціаніну. На відміну від інших фікобілінпротеїнів їхня молекула складається лише з одного типу білкових одиниць (молекулярна маса 15 кДа), причому кожна субодиниця несе одну молекулу ало-фікоціаніну.
Алофікоціанін характерний для синьозелених, хоча трапляється також у червоних і криптомонадових водоростей. А загалом поширення окремих типів пігментів фікобілінів серед синьозелених, червоних і бурих водоростей досить специфічне.
Слід зазначити, що на відміну від хлорофілів і каротиноїдів, локалізованих у ламелах, фікобіліни концентруються або в стромі, або формують особливі впорядковані ансамблі на поверхні мембран — фікобілісоми (рис. 10).
Як правило, в таких водоростях фікобіліни присутні в значно більшій концентрації порівняно з хлорофілами, тому саме вони і зумовлюють їхнє забарвлення. Фікоціаніни, фікоеритрини і алофікоціаніни трапляються в різних співвідношеннях, причому залежно від умов освітлення формується переважно такий набір пігментів, який найкраще використовує відповідний спектп.
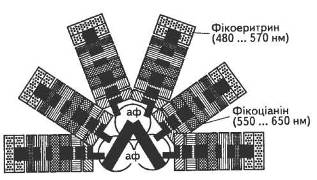
Рис. 10. Модель фікобілісоми: аф — алофікоціаніни
Пігменти даної групи виконують функцію світлозбиральної антени та забезпечують ефективну передачу поглинутої ними енергії сонячного світла до хлорофілів а.
Фікобіліни зумовлюють явище філогенетичної хроматичної адаптації водоростей в їхній вертикальній зональності.
Як відомо, червоні промені, що відповідають спектру поглинання хлорофілу, поглинаються, проходячи крізь товщу води. Вже на глибині 34 м зникають червоні промені, 177 м — жовті, 322 м — зелені, 500 м — сині та фіолетові. Відповідно до цього певні екологічні ніші займають спочатку зелені, потім синьо-зелені і, нарешті, червоні водорості. Наявність фікобілінів дає змогу водоростям у процесі фотосинтезу використовувати промені, які проникають на певну глибину, та займати відповідні екологічні ніші.
3.4 Оптичні властивості фотосинтетичних пігментів
Оптичні властивості пігментів зумовлені спільною для всіх структурою. Як основні, так і допоміжні пігменти характеризуються чітко вираженим чергуванням подвійних та одинарних зв'язків. Кожному зв'язку відповідають два електрони. Біля кожного атома вуглецю міститься вісім валентних електронів, які є спільними для даного атома вуглецю та для сусідніх його атомів. Подвійний зв'язок С=С формує пара електронів, які утворюють σ- та π-зв'язок. Крім того, є пара π-електронів, яка може переміщуватися по всьому вуглецевому ланцюжку (так званий резонанс). Завдяки резонансу молекула хлорофілу набуває додаткової стабільності. Спарені електрони пов'язані з усією системою спряжених подвійних зв'язків, а не з окремими атомами, тому вони легко збуджуються квантами світла. Енергія такого збудження настільки незначна, що легко забезпечується променями денного світла (380...720 нм).
Порфірини — найбільш стабільні та інертні органічні речовини. В порфіриновому ядрі хлорофілу система спряжених зв'язків замкнута й π-електрони можуть циркулювати, тобто характеризуються ще більшою свободою переміщення, ніж у каротиноїдах. Крім того, порівняно з каротиноїдами у хлорофілів є додаткові можливості для поглинання світла. Адже у кожного атома азоту три пари електронів спільні для сусідніх атомів вуглецю. На орбіталі є ще одна пара електронів, що спрямована до магнію. Це так звані л-електрони, які також здатні поглинати кванти.
3.5 Біосинтез пігментів фотосинтетичного апарату рослин
Фотосинтетичні пігменти — досить складні органічні сполуки, тому їх біосинтез охоплює низку етапів. Хлорофіли, як і подібні до них за хімічними властивостями цитохроми (Fe-порфірини), характеризуються єдиним процесом синтезу від простих складових до тетрапірольних структур (виразний приклад єдності шляхів метаболізму у тварин і рослин). Шляхом тетрапірольного синтезу утворюються й інші речовини, які використовують світло як сигнал (наприклад, фітохромна система), так що тетрапірольні структури формують майже весь залежний енергетичний та інформаційний апарат рослин.
Доведено, що в мембранах етіопластів і хлоропластів локалізовані специфічні центри біосинтезу пігментів, де й зосереджуються їхні попередники — протохлорофіліди та хлорофіліди. Реакції, що забезпечують синтез протохлорофіліду, відбуваються за відсутності світла, тоді як утворення з протохлорофіліду хлорофіл іду вимагає освітлення.
Біосинтез порфіринів у вищих рослин відбувається в три етапи:
• синтез амінолевулінової кислоти (АЛК) з п'ятивуглецевих дикарбонових (глутамінової, можливо, о-кетоглутарової) кислот;
• циклізація двох молекул АЛК з утворенням піролу порфобіліно-гена;
• утворення та циклізація тетрапіролу, формування магнійпротопорфірину, ферментативна етерифікація хлорофіліду фітолом.
У прокаріот і евкаріот АЛК синтезується із різних джерел: прокаріоти використовують для її синтезу гліцин і сукциніл-КоА, при цьому гліцин, перш за все, є джерелом азоту для побудови піроль-их кілець. Евкаріоти використовують різноманітні п'ятивуглецеві сполуки, зокрема глутамат, який спочатку фосфорилюється, а потім відновлюється.
На першому етапі синтез АЛК локалізований в хлоропластах, хоча самі ферменти, необхідні для цього, синтезуються в цитоплазмі клітини під контролем ядерних генів, а потім транспортуються в хлоропласти. Глутамінова кислота через 2-гідрооксиглута-рову перетворюється на 4,5-діоксивалеріанову (пентанову), яка потім амінується за рахунок аланіну чи інших амінокислот. Можливі й інші шляхи синтезу АЛК, які, ймовірно, локалізовані в різних компартментах клітини і по-різному регулюються. Здебільшого саме синтез АЛК обмежує швидкість біосинтезу хлорофілів, тому регуляція утворення даного проміжного продукту — надзвичайно важливий етап формування фотосинтетичного апарату рослин (рис. 11).
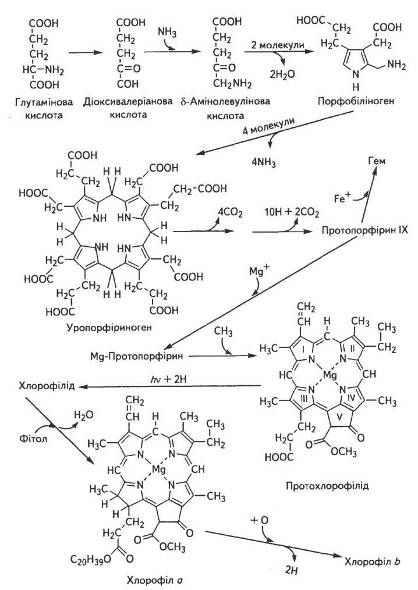
Рис. 11. Основні етапи біосинтезу хлорофілів
На другому етапі відбувається альдольна конденсація двох молекул АЛК з наступною ізомеризацією спочатку до піролу, а потім до порфобіліногену. Потім із чотирьох піролів синтезується уропорфіриноген, який поетапно перетворюється на протопорфірин IX.
На третьому етапі в результаті полімеризації чотирьох молекул порфобіліногена виникає тетрапірол, а пізніше, при замиканні кільця, утворюється уропорфіриноген І і ІІ. Перетворення порфобіліногена на уропорфіриноген ІІІ каталізується уропорфіриноген-1-синтетазою за наявності іншого білка КоА-синтетази.
Характерно, що в рослинній клітині лінійні тетрапіроли можуть бути хромофорними групами фітохрому, а циклічні — використовуватися для біосинтезу порфіринів, цитохромів, каталази чи пероксидази.
Уропорфіриноген III через серію реакцій окиснення та декарбоксилування перетворюється на протопорфірин IX. Завершальні реакції синтезу хлорофілів із протопорфірину IX повністю ще не з'ясовано, хоча відомо, що першою стадією, ймовірно, є ферментативне введення в молекулу атома магнію, потім метилування пропіонатного бічного ланцюга, насичення вінільної групи в кільці піролу IV, а відтак — замикання кільця V з утворенням протохлорофілу.
На цьому ж етапі формування молекули хлорофілу а відбувається етерифікація фітолом, який синтезується з ацетил-КоА через мевалонову кислоту.
Незважаючи на багато зусиль різних вчених, поки що нікому не вдаюся з'ясувати, здавалося б, просте питання: яким чином здійснюється біосинтез хлорофілу b. Цей хлорофіл, який завжди є в зелених листках, імовірно, утворюється з хлорофілу а заміщенням метильної групи СН3 біля другого пірольного кільця на альдегідну —СОН. Однак очевидний шлях — через окислення молекули хлорофілу а — поки що не доведено, а ніякого іншого шляху досі не знайдено.
Доведено, що швидкість синтезу та розпадання молекули хлорофілу приблизно однакова, тому його пул постійно оновлюється в хлоропласті. Нагадаємо, що в 1960 p. P. Вудворд і М. Штрель штучно синтезували молекули хлорофілу.
Важливе значення для синтезу хлорофілів має мінеральне живлення рослин. Для цього передусім необхідна достатня кількість заліза — каталізатора синтезу хлорофілів. Залізо необхідне для синтезу амінолевулінової кислоти та протопорфіринів. Для синтезу порфіринів має бути нормальне забезпечення азотом і магнієм, тому що обидва ці елементи входять до складу хлорофілу. Також потрібні йони міді, що забезпечують утворення стійких комплексів між хлорофілом і білком.
Біосинтез каротиноїдів розпочинається з ацетил-КоА через ме-валонову кислоту, гераніл-гераніолпірофосфат до лікопіну, який є попередником усіх інших каротиноїдів (рис. 12).
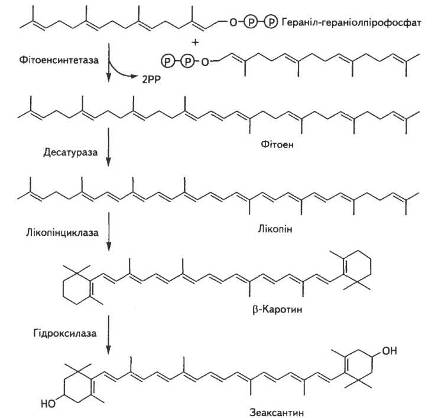
Рис. 12. Біосинтез каротиноїдів
Синтез їх відбувається в темряві, хоча різко прискорюється на світлі. Механізм біосинтезу фікобілінпротеїнів майже не вивчено.
Розділ 4. Енергетика фотосинтезу
4.1 Первинні процеси фотосинтезу
Фотосинтез — це насамперед фізична проблема. Первинні процеси його розпочинаються з миттєвих реакцій поглинання електромагнітної енергії сонячних квантів (τ = 1015 с), її запасання та стабілізації у вигляді електронного збудження, міграції енергії в пігмент-ліпопротеїдному комплексі хлоропластів:
![]()
Міграція — це процес передачі енергії електронно-збудженого стану від молекули пігменту, яка поглинула квант світла, до іншої молекули.
Ймовірний механізм енергетичного перенесення — міграція екситону, або індуктивний резонанс. Екситонний механізм міграції енергії характерний для гомогенних молекул пігментів.
Екситон — це елементарний стан електронного збудження, енергія якого делокалізована.
Екситон — це квазічастинка, яка існує ~ 10-5с і характеризується певною енергією та імпульсом. Електронно-збуджений стан мігрує як спін-спряжена пара електрон—дірка. Рух екситону здійснюється як дискретне (перескоки), або як неперервне (когерентне) зміщення.
Індуктивний резонанс забезпечує перенесення енергії без випромінювання фотонів у результаті диполь-дипольної взаємодії молекул донора і акцептора, наприклад між хлорофілами а і b. Швидкість перенесення енергії залежить від величини відстані між цими молекулами та ступеня перекривання спектра флуоресценції донора зі спектром поглинання акцептора.
Передачу енергії крізь молекули різних пігментів визначають як гетерогенне перенесення, тоді як аналогічний процес між гомологічними молекулами називають гомогенним перенесенням. Дослідження флуоресценції є доказом можливої передачі енергії гетерогенного або гомогенного типу. Так, у разі опромінення пігментної системи монохроматичним світлом, наприклад таким, що поглинається тільки хлорофілом b, можна зафіксувати спектр флуоресценції хлорофілу а.
Вдалося виявити міграцію енергії між такими парами: каротиноїди → фікоеритрин → фікоціанін → хлорофіл b → хлорофіл a; хлорофіл а → хлорофіл а. Завжди можлива міграція енергії від кожного з пігментів безпосередньо на хлорофіл а.
Енергія завжди мігрує у напрямі від пігментів з максимумом поглинання в зоні коротших хвиль до молекул, максимум поглинання яких розміщений у довгохвильовій зоні.
Нині чітко доведено гетерогенність хлорофілів у мембрані, тобто спектральна дискретність хімічно тотожних молекул пігментів, яка зумовлена відмінностями в характері взаємодії пігменту з навколишнім середовищем.
Як відомо, світло має водночас хвильові й корпускулярні властивості, тобто поширюється як хвиля та поглинається квантами, або фотонами. Фотоактивні речовини, в тому числі пігменти, здатні поглинати кванти світла. Це означає, що незбуджена молекула хлорофілу перебуває в тепловій рівновазі з навколишнім середовищем, і такий стан називають основним (So). У такому стані валентні електрони займають найнижчі енергетичні рівні (причому, згідно з принципом Паулі, не більше двох електронів із антипаралельними спінами на кожній орбіталі). Сумарний спін (вектор магнітних моментів) усіх електронів молекули дорівнює нулю. У разі поглинання кванта світла молекула хлорофілу переходить на один зі збуджених коливальних підрівнів електронного рівня, тобто на вищу вільну орбіталь. Такий перехід не змінює напряму електронного спіну й відбувається за 1015 с. Якщо в збудженому електроні зберігається той самий напрям спіну, то молекула перебуває в так званому синглетному стані (рис. 13).
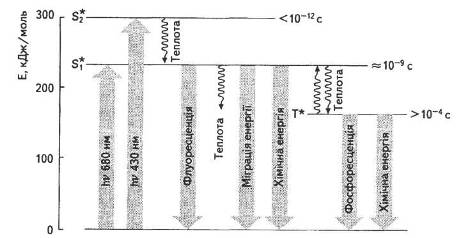
Рис. 13. Енергетичні стани молекули хлорофілу
Якщо під час переходу на вищу орбіталь спін змінює свій напрям, таке збудження називають триплетним (спіни стають паралельними), при цьому сумарний спін дорівнює одиниці.
Молекула хлорофілу може поглинати світло як у червоній, так і в синій частині спектра, тобто одержувати різну кількість енергії, а отже, переходити на перший або другий синглетний рівні. Час існування збудженої молекули незначний (на першому синглетному рівні — 10-8, 10-9 с, а на другому — 10-12 с), після чого вона повертається на вихідний енергетичний рівень. Відбувається це різними шляхами (рис. 14).
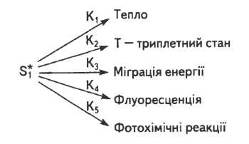
Рис. 14. Можливі шляхи трансформації енергії першого збудженого синглетного стану хлорофілу
Первинна фотофізична стадія розпочинається з поглинання світла пігментами. Кожна молекула хлорофілу поглинає 4...40 квантів світла за секунду. Міграція енергії забезпечує ефективне збирання квантів світла антенним світлозбиральним комплексом (СЗК) і транспортування їх до молекул хлорофілу, що поєднані з фотохімічними реакціями.
4.2 Фотосинтетична одиниця, реакційний центр, фотосистема
Молекули хлорофілу, які перетворюють сонячну енергію, локалізуються приблизно в 1000 тилакоїдів кожного хлоропласту таким чином, що за відповідного освітлення одиницею поглинання є переважна їх більшість. Водночас вони є своєрідним «озером», у якому плаває множина так званих фотосинтетичних одиниць.
Уявлення про фотосинтетичну одиницю (ФСО) як комплекс пігментів, що бере участь у поглинанні, збиранні квантів і передачі їх спеціалізованим молекулам хлорофілу, майже одночасно було сформульовано Дж. Гафроном, Р. Емерсоном і У. Арнольді. На їхню думку, основна маса молекул пігментів, які складають ФСО, забезпечує збирання енергії та передачі її спеціальним компонентам, що фотохімічно перетворюють цю енергію. Таким чином вони виконують функцію пускового механізму фотосинтезу (рис. 15).
Такий функціонально активний комплекс було названо реакційним центром (РІД). Фотохімічні реакції здійснюють лише пігменти РІД, тоді як усі інші лише поглинають світло і переносять енергію збудження на хлорофіли реакційних центрів. Згодом було доведено, що один РІД припадає приблизно на 400...600 молекул пігменту. Частка пігментів РІД становить близько 1 % від суми всіх пігментів хлоропластів.
Загальноприйняті нині уявлення про фотохімічні реакції хлорофілів РІД сформувалися наприкінці 50-х років минулого століття під впливом класичних робіт Р. Емеосона зі співробітниками. Вони експериментально довели, що в процесі фотосинтезу рослин беруть участь дві різні фотохімічні реакції. Як виявилося, швидкість фотосинтезу за спільної дії квантів червоного світла (700 нм) та короткохвильового (680 нм) була більшою, ніж сума швидкостей фотосинтезу в разі окремого освітлення цими квантами (ефект підсилення Емерсона).
Зокрема, квантовий вихід фотосинтезу (максимальне число О2, які звільняються в перерахунку на кожний поглинутий квант) неадитивно зростав, інколи аж до 40 %. Такий ефект підсилення було пояснено функціонуванням двох фотохімічних систем (ФС І та ФС II), які послідовно виконують первинні фотохімічні реакції. Нині навіть вдалося розділити фотосистеми як за допомогою інгібіторів фотосинтезу (хімічне розділення), так і диференційним центрифугуванням (механічне розділення). Тилакоїдну мембрану вдалося розділити на дві фракції — легку та важку, які осаджувалися відповідно при 24 000 та 144 000 д. До легкої входять в основному фрагменти мембран тилакоїдів строми, які є складовими частками ФС І. Аналіз важкої фракції показав, що вона збагачена компонентами ФС II.
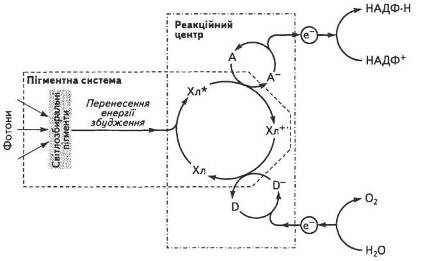
Рис. 15. Спрощена схема роботи пігментної системи хлоропласта
Згідно з сучасними уявленнями, хлорофіли реакційних центрів виконують основну функцію в перетворенні енергії сонячних квантів. Завдяки специфічним зв'язкам та організації їхніх молекул-донорів і молекул-акцепторів хлорофіли здатні використовувати одержану енергію збудження для утворення потоку електронів шляхом оборотних окисно-відновних реакцій. В результаті цього в тилакоїдній мембрані формується протонний і електричний градієнти. Максимуми поглинання хлорофілів реакційних центрів двох систем, за яких відбуваються окисно-відновні процеси, різні: максимум хлорофілу реакційного центру ФС І (Р700) — 700 нм, тоді як хлорофілу реакційного центру ФС II (Р680) — 680 нм. Кожен із хлорофілів має відповідні донори та акцептори електронів, які започатковують серію окисно-відновних перетворень відповідних переносників електронів у тилакоїдній мембрані. Ймовірна схема структури реакційних центрів ФС І і ФС II наведена на рис. 16.
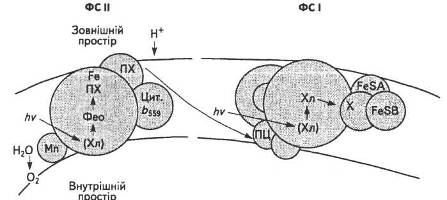
Рис. 16. Реакційні центри ФС І і ФС II
Донори електронів локалізовані всередині, тоді як їхні акцептори — зовні тилакоїдної мембрани, що і забезпечує миттєвий розподіл заряду на мембрані.
Фотосинтетична одиниця (ФСО) — це мінімальне функціональне угруповання тилакоїда, здатне до фотосинтетичного розподілу заряду і складається з пігментів антенного комплексу та реакційних центрів.
За останніми даними, на один хлоропласт припадає 2 • 106 ФСО. Ці найдрібніші одиниці здатні здійснювати окиснення й відновлення НАДФ+, мають антенні комплекси пігментів-світлозбирачів, двіфотосистеми, що формують разом із багатьма переносниками електронно-транспортний ланцюг (ЕТЛ) тилакоїдної мембрани.
Досі немає єдиної думки щодо структури ФСО. Насамперед, було запропоновано моноцентральну модель ФСО, згідно з якою в усьому комплексі є лише один РІД. Більшість учених віддають перевагу мультицентральній моделі, яка припускає наявність кількох реакційних центрів (рис. 17).
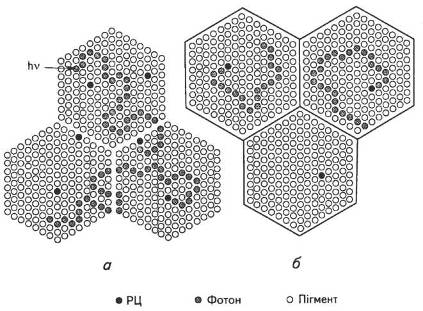
Рис. 17. Модель мультииентральної (а) та уніцентральної (б) фотосинтетичних одиниць:
о — пігмент антенного комплексу;
• — Р70О або Р680 в реакційному центрі;
* — молекула пігменту антени, яка перейшла у фотозбуджений стан після поглинання фотона або внаслідок перенесення енергії
В хлоропластах вищих рослин існує як мінімум два типи фото-;истем: ФС І і ФС II.
Фотосистема — це дискретна одиниця організації в тилакоїдах певних пігментів разом з іншими молекулами — переносниками електронів і протонів, які беруть участь у світловій стадії фотосинтезу.
Точна локалізація фотосистем у тилакоїдній мембрані хлоропластів поки що не відома. Вважають, що мембрани тилакоїдів строми містять переважно ФС І, а мембрани гран — ФС І і ФС II. Таким чином, у тилакоїдній мембрані молекули пігментів і переносники електронів об'єднані в надмолекулярні функціональні комплекси.
4.3 Z-cxeмa фотосинтезу
Р. Хілл і Ф. Бендалл запропонували схему електронної стадії фотосинтетичного процесу, яка після низки уточнень одержала назву Z-схеми фотосинтезу (рис. 18).
Переносники електронів ЕТЛ розміщуються відповідно до зменшення електровід'ємного редокс-потенціалу. В фотосинтезі вищих рослин перенесення електрона від двох молекул води (окисно-відновний потенціал 0,82 В) до НАДФ+ (окисно-відновний потенціал — 0,32 В) здійснюється за рахунок енергії чотирьох квантів світла з участю відповідно двох фотосистем, що діють послідовно. Механізм передачі електронів в обох фотосистемах — одноелектронний.
Під час поглинання кванта світла ФС II за період, менший за 105 с, електрон від води переноситься в ЕТЛ, проти термодинамічного потенціалу, а саме: від позитивного редокс-потенціалу фередоксину на НАДФ+. Саме в цьому полягає основна суть фотосинтезу — адже в процесі такого транспортування запасається вільна енергія поглинутих квантів. У мембрані тилакоїду розміщується близько 200 таких ланцюгів.
Шлях електрона від води до НАДФ+ з участю обох фотосистем називають нециклічним.
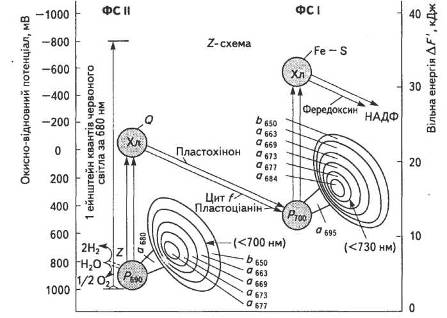
Рис. 18. Z-схема фотосинтезу
Існує й циклічне транспортування електронів з участю лише ФС І, яке надходить від неідентифікованого компонента X на фередоксин, далі — на цитохром Ь6 (Ео = — 0,18 В) і пластохінон (Пх). З відновленого Пх • Н2 електрони транспортуються на цитохром f, далі — на пластоціанін і, нарешті, заповнюють дірку того самого пігменту Р700.
Можливий також псевдоциклічний шлях транспортування електрона, коли в результаті передачі електрона на кисень він відновлюється до Н2О, чим практично маскується процес фотолізу води в ФС II (рис. 19).
Як видно зі Z-схеми, на шляху від однієї молекули Н2О до НАДФ+ крізь переносники передаються два електрони. Якщо один квант енергії, що надходить в кожну фотосистему, піднімає на вищий рівень лише один електрон, то для перенесення двох електронів треба мінімум чотири кванти. Світлозбиральний апарат, який постачає енергію, складається з лабільного світлозбпрального комплексу (СЗК), що обслуговує обидві фотосистеми, та пігмент-білкових антенних комплексів ФС І і ФС II. Уявлення про Z-схему фотосинтезу на цей час є загальноприйнятим, хоча постійно піддається певним змінам і доповненням.
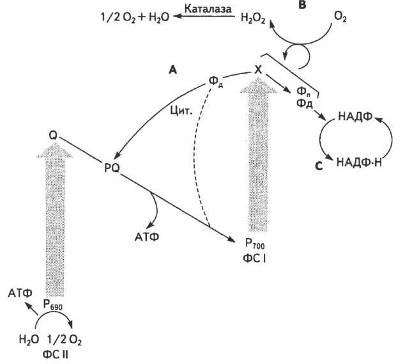
Рис. 19. Можливі шляхи перенесення електронів у хлоропластах:
А — циклічний; В — псевдоциклічний; С — нециклічний
Висновки
Особливості морфології, анатомії та фізіології листка повною мірою забезпечують здійснення його основної функції — фотосинтезу. Функціонування продихів регулює газообмін — надходження вуглекислого газу, виділення кисню та випаровування води.
Фотосинтетичні пігменти — досить складні органічні сполуки, тому їх біосинтез охоплює низку етапів. Хлорофіли, як і подібні до них за хімічними властивостями цитохроми (Fe-порфірини), характеризуються єдиним процесом синтезу від простих складових до тетрапірольних структур.
Пігменти пластид відносять до трьох класів, а саме: хлорофіли, каротиноїди та фікобіліни. Найважливіше значення в процесі фотосинтезу належить зеленим пігментам — хлорофілам. Нині відомо близько десяти хлорофілів, які відрізняються хімічним складом, забарвленням і поширенням серед живих організмів. Основними пігментами, без яких фотосинтез не відбувається, є хлорофіл а для зелених рослин і бактеріохлорофіл для фототрофних бактерій. Пігменти — це сполуки, які вибірково поглинають світло у видимій (400...700 нм) частині спектра.
Непоглинені ділянки сонячного спектра відбиваються, що й зумовлює забарвлення пігменту. Зелений пігмент хлорофіл поглинає червоні та сині промені, тоді як зелені в основному відбиваються.
Фотосинтез — це насамперед фізична проблема. Первинні процеси його розпочинаються з миттєвих реакцій поглинання електромагнітної енергії сонячних квантів, її запасання та стабілізації у вигляді електронного збудження, міграції енергії в пігмент-ліпопротеїдному комплексі хлоропластів.
Світлоіндуковане векторне транспортування електронів між двома фотосистемами, яке розпочинається від води й закінчується НАДФ+, забезпечується завдяки специфічному й несиметричному розташуванню низки окисно-відновних компонентів тилакоїдної мембрани — переносників електронів і протонів, які в сукупності складають електронно-транспортний ланцюг.
Список літератури
1. Бугай С.М. Растенееводство. – К.: Вища школа, 1975. – 375 с.
2. Избранные главы физиологии растений / В. Ф. Гавриленко, М. В. Гусев, К. А. Никитина. — М.: Изд-во Моск. ун-та, І986. — 436 с.
3. Кочубей С. М. Организация фотосинтетического аппарата высших растений. — Киев: Альтерпрес, 2001. — 204 с.
4. Мокроносов А. Т., Гавриленко В. Ф. Фотосинтез: физиологические и биохимические аспекты. — М.: Изд-во Моск. ун-та, 1992. — 319 с.
5. Максимов Н.А. Краткий курс физиологии растений, 1958.
6. Мусієнко М. М. Фотосинтез. — Киев: Вища школа, 1995. — 247 с.
7. Мусієнко М.М. Фізіологія рослин. – К.: Либідь, 2005. – 808 с.
8. Рубин Б.А. Курс физиологии растений. – М.: Высшая школа, 1961. –583 с.
9. Силаева А. Й. Структура хлоропластов и факторы среды. — Киев: Наук, думка, 1978. — 157 с.
10. Фотосинтез: В 2 т. / Под ред. Р. Говинджи. — М.: Мир, 1987. — Т. 1. 728 с; Т. 2. 460 с.
11. Эдварде Дж., Уокер Д. Фотосинтез С3- и С4-растений: механизмы и регуляция. — М.: Мир, 1986. — 590 с.