
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Рефераты по косметологии
Рефераты по криминалистике
Рефераты по криминологии
Рефераты по науке и технике
Рефераты по кулинарии
Рефераты по культурологии
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Рефераты по косметологии
Рефераты по криминалистике
Рефераты по криминологии
Рефераты по науке и технике
Рефераты по кулинарии
Рефераты по культурологии
Курсовая работа: Анатомические особенности строения психрофитных растений семейства вересковых
Курсовая работа: Анатомические особенности строения психрофитных растений семейства вересковых
ОГЛАВЛЕНИЕ
Введение
Глава I. Характеристика физико-географических условий Красноселькупского района Ямало-ненецкого автономного округа (ЯНАО).
1.1 Геологическое строение и рельеф, геокриологические условия
1.2 Климат
1.3 Почвенный покров
1.4 Растительность
Глава II. Экология и биология психрофитных растений семейства вересковые
2.1 Особенности биологии психрофитных растений
2.2 Обзор видового состава вересковых Красноселькупского района
Экспериментальная часть
Глава III. Морфо-анатомические и экологические особенности строения психрофитных растений семейства вересковые
3.1 Материалы и методы исследования
3.2 Анатомические особенности строения побегов вечнозеленых и листопадных психрофитных кустарничков семейства вересковые
Заключение
Библиография
Приложение
ВВЕДЕНИЕ
Специфика природных условий Севера и, в частности, экстремальные значения абиотических факторов, такие, как непрерывное освещение в период вегетации, слабая интенсивность света, обусловленная широтой и усугубленная постоянной облачностью и туманами, не могли не сказаться на морфологических, эколого-физиологических и биологических признаках, свойствах и особенностях северных растений. По совокупности данных показателей выделили отдельную экологическую группу холодостойких растений – это растения психрофиты.
К ним относят виды, произрастающие в тайге, тундре, высокогорьях. Сюда же могут относиться растения, живущие в условиях холодного, плохо прогревающегося даже летом, сфагнового болота.
Среди психрофитных растений встречаются представители таких семейств: осоковые, гвоздичные, крестоцветные, сложноцветные, большое количество видов семейства вересковые
Растения тундры живут в очень суровых условиях, что накладывает особый отпечаток на их облик. Большинство выше названных растений обитают в нашем районе и являются удобным объектом для наблюдений и исследований.
Необходимость экологизации школьного биологического образования в настоящий момент ни у кого не вызывает сомнения – ведь и возникновение, и решение экологических проблем тесно связано с биологией, ее прикладными и теоретическими аспектами.
Задачей же школьного курса биологии является, прежде всего, формирование диалектико-материалистических взглядов на природу и на взаимодействие общества и природы; овладение учащимися знаниями и умениями для рационального использования природных ресурсов и охраны окружающей среды, оценки природной и хозяйственной обстановки в своей местности, воспитание норм и правил поведения в природе.
Все вышесказанное и обуславливает актуальность выбранной темы исследования.
Цель исследования:
- изучить некоторые анатомические особенности строения психрофитных растений семейства вересковых;
Объект исследования: психрофитные растения семейства вересковые.
Гипотеза: психрофитные растения имеют ряд морфо-анатомических особенностей, связанных с условиями их произрастания. Исходя из целей, объекта, предмета и гипотезы исследования необходимо решить следующие задачи:
1. Проанализировать литературу и обобщить методический опыт по проблеме исследования.
2. Выяснить какие растения семейства вересковые относятся к группе психрофитных растений. Изучить их экологию и биологию.
3. Изучить анатомические особенности строения стеблей и листьев некоторых психрофитных растений семейства вересковые.
Глава I. Характеристика физико-географических условий Красноселькупского района Ямало-ненецкого автономного округа (ЯНАО)
На северо-востоке Западно-Сибирской равнины, в бассейне крупной реки Таз, располагается один из самых примечательных районов Ямало-Ненецкого автономного округа (ЯНАО) – Красноселькупский. Уникальность его положения в том, что почти в течение столетия здесь находился географический центр нашего государства: еще в начале XX века уроженец Тобольской губернии великий ученый Д.И. Менделеев определил, что географический центр Российской империи находится в верховьях реки Таз. Позже, на территории Красноселькупского района, располагался центр Советского Союза, в честь этого 24 августа 1983 года на правом берегу Таза, в полутора километрах ниже реки Малая Ширта, был установлен памятный знак.
Правда, в настоящее время географический центр Российского государства сместился к востоку – в центральные районы Красноярского края. Сейчас можно отметить другую уникальность Красноселькупского района – это одно из немногих мест Тюменского Севера, которое еще слабо затронуто процессом нефтегазового освоения и в значительной степени сохранило первозданную природу и традиционный образ жизни коренного населения.
Красноселькупский район расположен в юго-восточной части ЯНАО. С севера на юг он протянулся на 500 километров, с запада на восток – более чем на 350 километров. Площадь района – 106,8 тыс.кв.км., что составляет 14,2% территории ЯНАО.
Рождение Красноселькупского района совпало с появлением на карте страны Тюменской области. Он образован Указом Президиума Верховного Совета РСФСР 10 августа 1944 года в ЯНАО
1.1 Геологическое строение и рельеф, геокриологические условия
Геологическая история Западно-Сибирской равнины, на территории которой размещается Красноселькупский район, до недавнего времени представлялась относительно простой. Это гигантская чаша, дно и стенки которой состоят из горных пород, образовавшихся 500-250 млн лет назад. Ее глубина достигает 4-5 км, и только на юге в районе Верхнее-Тазовской возвышенности она равна 2,0-2,5 км. Дно представляет собой складчатый фундамент Западно-Сибирской платформы. Она образована, в основном, магнетическими (изверженными) и метаморфическими (преобразованными) породами. Этот платформенный чехол сформирован гигантским циклом осадконакопления. Он начался в континентальных условиях, продолжился в морских и заканчивается снова в континентальных. Преобладают озерно-аллювиальные песчано-глинистые и озерные, преимущественно глинистые, отложения. Однако на значительных площадях можно встретить валуны и гальку. Невольно возникает вопрос: какая же сила оказалась способной перенести камни на практически ровной поверхности? Такой силой, по мнению ученых явился лед. Глобальные похолодания, захватившие Землю в конце неогена (700тыс. лет назад) закончились только около 10 тыс. лет назад. В этот период лед то наступал, покрывая всю современную территорию района, то отступал, оставляя на поверхности свои следы – валуны, гальку и тому подобное. От ледникового периода на больших пространствах еще сохранилась масса межморенных остаточных приледниковых озер, а в южной части – термокарстовых и просадочных озер. Современный рельеф территории Красноселькупского района обусловлен геологическим развитием, тектоническим строением и влиянием различных экзогенных (поверхностных) рельефообразующих процессов. Основные орографические (рельефные) элементы находятся в тесной зависимости от тектонического строения плиты, перекрытой мощным чехлом рыхлых отложений. Хотя слои в чехле залегают почти горизонтально, тектонические движения местами нарушили эту горизонтальность и образовали огромные впадины и своды, в которых обычно накапливались нефть и газ. На поверхности рельеф местности осложнен небольшими поднятиями (возвышенностями). Но все-таки низменные территории преобладают. Вот здесь-то и встречаются в изобилии знаменитые сибирские болота, многочисленные озера и извилистые реки.
Основные микроформы рельефа – бугры пучения и термокарстовые озера связаны с широким распространением многолетней мерзлоты, которая оказывает решающую роль в формировании современного рельефа.
Мерзлотные формы рельефа распространены повсеместно – от междуречий до пойм – и проявляются в морозобойном трещинообразовании, пучении и термокарсте. Процесс трещинообразования преобладает в торфяниках и минеральных грунтах, где имеются линзы и прослои льда в сочетании с низкими температурами. Данный процесс чаще наблюдается на возвышенных участках, с которых сдувается снег, в результате чего талая вода быстро замерзает в трещинах, разрушая почву и образуя полигональные формы. Полигоны имеют формы четырех-пяти- и шестиугольников, покрытых мхами и лишайниками и окаймленных кустарничками.
В бассейне Таза широко распространены торфяники. Их образование связано с процессами морозобойного растрескивания и пучения. Причем бугристые торфяники не однородны: выделяются плоско- и крупнобугристые. Плоскобугристые торфяники господствуют в лесотундре на водораздельных пространствах. Высота торфяников 2-3 м., склоны пологие, реже крутые.
Для северной тайги более характерны крупнобугристые торфяники, с максимальной высотой до 10 метров. Среди плоско- и крупнобугристых торфяников много термокарстовых озер. Другу от друга они часто "отгорожены" узкой бровкой торфа с линзами льда, протаивание которых приводит к спуску воды из озера. На дне спущенных озерных котловин талые отложения подвергаются промерзанию и пучению с образованием булгунняхов. Такие формы встречаются в бассейне Таза практически повсеместно, кроме поймы.
Нередки случаи заторфовывания термокарстовых озер. На их месте образуются заболоченные понижения – хасыреи. Кроме хасыреев типичной формой рельефа являются и аласы (поляна, луг, открытая небольшая равнина). Аласовый рельеф развит обычно вблизи истоков рек. Аласы имеют глубину до десяти-двадцати метров, на дне располагаются сточные, заросшие по берегам озера.
Все сказанное подтверждает, что поверхность территории, на которой размещается Красноселькупский район, довольно сложна, а процесс ее формирования и преобразования не закончился.
Суровый климат и отрицательные среднегодовые температуры способствовали широкому развитию на территории района многолетней ("вечной") мерзлоты. "Вечная мерзлота" - это мерзлые толщи горных пород, неоттаивающие на протяжении длительного времени – от нескольких лет до десятков и сотен тысяч лет. Мерзлотному состоянию способствует отрицательная температура воздуха в течение года.
Вечная мерзлота начала формироваться в конце неогена – начале четвертичного периода (около 700 тысяч лет назад), а ко времени максимального оледенения (10 000лет назад) она была распространена уже на всей территории Красноселькупского района. И, несмотря на то, что ледниковая эпоха давно закончилась, вечная мерзлота здесь в том или ином виде сохранилась, хотя характер ее залегания неодинаков.
В пределах Красноселькупского района выделяются две крупные мерзлотные области, последовательно сменяющие одна другую с севера на юг и различающиеся между собой характером залегания:
1. Слитного залегания современной и древней вечной мерзлоты.
Область слитного залегания современной и древней вечной мерзлоты распространена на севере района;
2. Разобщенного залегания современной и древней вечной мерзлоты. Южнее, в районе северной и средней тайги в виде "перелетков", или островов. Перелеток – сохранившийся в течение года слой мерзлотного грунта среди оттаявшего массива многолетней мерзлоты.
|
|
Вечная мерзлота оказывает большое влияние на хозяйственную деятельность человека. При сооружении жилых зданий, промышленных предприятий, железных и автомобильных дорог, трубопроводов изменяется термический режим пород, что нередко приводит к деформации фундаментов и даже разрушению зданий. Мощная толща льда деформируют обсадные колоны труб, глушит газовые фонтаны, сгущает нефть. При закладке в этих местах зданий специалисты изучают мерзлые грунты, учитывают их физические свойства, чтобы нейтрализовать неблагоприятные последствия.
1.2 Климат
Располагаясь на севере материка Евразия вблизи Северного Ледовитого океана и испытывая периодические воздействия воздушных масс, приходящих с Атлантического океана и Восточной Сибири, территория Красноселькупского района имеет континентальный климат. Это проявляется, в первую очередь, в большой разнице (амплитуде) температур самого теплого и холодного времени года. Она нарастает с юго-запада на северо-восток, где смягчающее влияние Атлантики уменьшается.
Между севером (Сидоровск) и югом (Толька) наблюдаются ощутимые погодные различия, обусловленные неодинаковым режимом солнечной радиации, неоднородностью подстилающей поверхности в холодное и теплое время года и характером циркуляции атмосферы. На температурный режим самое непосредственное влияние оказывает продолжительность солнечного сияния, связанная с углом наклона солнечных лучей и зависящая от географической широты.
Поступление солнечной радиации на территорию района неравномерно, что в первую очередь обусловлено наличием полярного дня и полярной ночи. В декабре-январе севернее Полярного круга длится полярная ночь, а с июня по июль – полярный1 день
Благодаря продолжительному освещению весной и летом длительность солнечного сияния по району колеблется незначительно: в апреле от 205 на севере района до 210 часов на юге, в июле, соответственно, от 295 до 330 часов. В среднем за год продолжительность солнечного сияния изменяется от 1500 часов на севере района до 1650 часов на юге. Уменьшение продолжительности солнечного сияния в лесотундре обусловлено частой облачностью (около 50 % облачных дней в году) и туманами. Наименьшая продолжительность солнечного сияния здесь отмечается зимой.
В течение семи-восьми месяцев температура воздуха отрицательная, поэтому среднегодовая температура воздуха на всей территории района также имеет отрицательные значения: от- 8,5˚С на юге, до – 6,1˚С на севере. Причем суровость зимы нарастает не с юга на север, а с юго-запада на северо-восток. Меридиональный ход изотерм выражен с ноября по март, что свидетельствует о преобладании в это время западного переноса воздушных масс сопровождающего циклонами, приходящими с северной Атлантики. В холодное время года западная часть района примерно на 1-2 градуса теплее по сравнению с восточной. Средние температуры самого холодного месяца – января – составляют – 27˚С на севере и –24˚С на юге, а в отдельные дни температура опускается ночью до – 60˚С и ниже. Абсолютный минимум, зарегистрированный на территории района, составил – 63˚С.
Изотермы летних месяцев идут почти широтно, что связано с преобладанием северных воздушных потоков. Такой ход изотерм наблюдается с июля по сентябрь. Летом в пределах района в отдельные периоды возможны жаркие дни. Абсолютный максимум температуры воздуха и на севере и на юге зарегистрирован на отметке +37˚С.
Продолжительная и холодная зима на территории Красноселькупского района благоприятствует накоплению снега и образованию устойчивого снежного покрова, что происходит почти одновременно на всей территории в первой половине октября, а разрушение снежного покрова на юге района начинается примерно на 12-15 дней раньше, чем на севере. Число дней со снежным покровом колеблется от 205 на юге до 225 на севере.
1.3 Почвенный покров
Почвенный покров Красноселькупского района не отличается большим разнообразием. Он сложился в условиях недостатка тепла, избыточного увлажнения и близкого залегания мерзлоты. При движении с севера на юг наблюдается его мозаичность, вызванная различием минералогического и механического состава почвообразующих пород и увеличением термических ресурсов.
По происхождению и экологическим свойствам почвенный покров делится на две большие группы:
1. Почвы водоразделов, или зональные.
К зональным типам относятся – подзолистые и болотно-подзолистые песчаные, таежно-поверхностно-глеевые, таежные заболоченные и болотные почвы.
Почвы подзолистого типа представлены маломощными лесотундровыми глеевато-слабоподзолистыми и слабоподзолистыми иллювиально-железистыми.
Лесотундрово-глеевато-слабоподзолистые почвы развиты в области ледниковых увалов и в центральных частях надпойменных террас. Они формируются под ерником и тундровым редколесьем, имеют серовато-лиловый цвет, малое содержание гумуса. Эти почвы имеют кислую реакцию почвенной среды, бедны подвешенными формами фосфора и калия.
Слабоподзолистые иллювиально-железистые почвы приурочены к массивам песчаных отложений и лиственничных редколесий с глубоко оттаивающей мерзлотой.
В сельскохозяйственном (земледельческом) отношении почвы лесотундры малоперспективны, особенно иллювиально-железистые, отличающиеся наиболее низким потенциальным плодородием из-за адсорбции фосфора железом и быстрой потери элементов питания в результате высокой водопроницаемости почвенной толщи. Их окультуривание связано с большими затратами на удобрение.
В северной и средней тайге выделяются три типа наиболее распространенных почв: таежно-поверхностно-глеевые, подзолистые и болотные. Все они развиваются на водоразделах при глубоко залегающих грунтовых водах: первые – на глинистом и суглинистом, вторые – на супесчаном субстрате. Таежно-поверхностные глееватые почвы приурочены к плоским элементам рельефа и лиственнично-еловым редколесьем с кустарничковым покровом.
На пониженных элементах рельефа – в западинах, по периферии болотных массивов с близким (до 1метра) уровнем залегания грунтовых вод расположены иллювиально-гумусовые подзолы. Эти почвы имеют контрастную окраску – под светлым подзолистым горизонтом располагается кофейно-коричневый горизонт, обогащенный гумусом (до 2%) и окислами алюминия (до 4%). Они используются при очаговом земледелии (овощной тип), но для получения стабильных урожаев нуждаются в улучшении водно-теплового режима, требуют внесения извести и органо-минеральных удобрений.
Особую группу образуют остаточно-торфянистые почвы бугров пучения и плоскобугристых торфяников. Образованию данного типа почв благоприятствует высокая переувлажненность территории с низкими скоростями разложения органических остатков болотной растительности и неглубокое залегание многолетнемерзлых пород. В сельскохозяйственном отношении эти почвы интереса пока не представляют.
2. Почвы речных долин, или интразональные.
К интрозанальным типам относятся аллювиальные
В Тазовской долинной провинции почвенный покров образован исключительно интразональными почвами – луговыми и подзолисто-иллювиально-гумусовыми. Благодаря тому, что река Таз и ее многочисленные притоки берут свое начало в относительно "богатых"
минеральными веществами районах, северные отрезки речных долин вместе с весенним половодьем получают дополнительное количество органо-минеральных веществ. Это обстоятельство делает почвы речных долин наиболее продуктивными и наиболее перспективными для сельскохозяйственного освоения.
1.4 Растительность
Суровые климатические условия, специфика почвогрунтов (бедность питательными веществами, нередко мерзлый характер грунтов, острый дефицит кальция, калия, фосфора в корнеобитаемом слое) определили сравнительную бедность флористического комплекса Красноселькупского района.
Растительный покров района формируют леса, болота и луга различных типов. Лиственничные редколесья на северных пределах района приурочены к речным долинам, наиболее хорошо дренируемым участкам склонов, надпойменным террасам. Почти везде в редколесьях встречаются вкрапления березы и ели.
В понижениях на водоразделах и в долинах развиты верховые и низинные болота, пушицевые кочкарники с моховым или мохово-лишайниковым покровом. Широко распространены растительные комплексы спущенных озер – хасыреев. Такие участки являются лучшими оленьими пастбищами летне-осеннего сезона. Русла рек окаймлены злаково-разнотравными ивняками.
На хорошо дренированных песчаных террасах развиты сосновые лишайниковые леса. Древостой их несколько разреженный, образован сосной с незначительной примесью березы бородавчатой и ели сибирской.
В речных долинах растительность более разнообразна и резко изменяется в зависимости от местоположения. В прирусловой части, на песках встречаются пойменные разнотравно-злаковые луга. Наиболее повышенные участки центральной поймы занимают еловые зеленомощные леса. Среди смешанных долинных лесов в прирусловой части поймы попадаются единичные крупные плодоносящие деревья кедра сибирского.
В долинах малых рек развиты кедровые и сосновые леса, в местах с обильным увлажнением и хорошим дренажем – березовые леса с участием ели сибирской, осины обыкновенной и пихты сибирской. Характерны пойменные ивняки, вытянутые узкой полосой вдоль речного русла. Они сменяются пойменными березовыми лесами с богатым кустарничковым подлеском и разнотравно-злаковым травяным покровом.
Грядово-мочажинные болота занимают почти половину всей заболоченной площади водоразделов. В надпочвенном покрове часто господствуют зеленые мхи, а в травяно-кустарничковом ярусе нередко обильна брусника, местами – черника.
Глава II. Экология и биология психрофитных растений семейства вересковые
Приспособление северных растений к относительно низким температурам носит как физиологический, так и морфологический характер. Многие виды образуют специфические жизненные формы – подушковидные, простратностелющиеся, розеточные и некоторые другие. Другим важным приспособлением является опушение различной степени и характера, препятствующее тепловому излучению и, следовательно, повышающее температуру опушенных органов, особенно в солнечные дни.
Успешное существование растений в суровых климатических условиях невозможно без ряда приспособлений к довольно короткому вегетационному периоду. Одно из них – быстрое начало роста побегов весной, несмотря на низкую температуру воздуха и почвы. Интенсивный рост в весеннее время возможен благодаря быстрой мобилизации и перемещению запасных углеводов из подземных частей растений в развивающие побеги и листья.
Другой особенностью является сформированность побегов будущего года в побегах возобновления уже к наступлению зимнего периода.
Из выше сказанного следует еще одна важная черта биоморфологической структуры психрофитных растений, а точнее спектра их жизненных форм – практически полное отсутствие однолетников (Алексеев Ю.Е., 2003).
2.1 Особенности биологии психрофитных растений
К группе психрофитных относят растения, произрастающие в тайге, тундре, на высокогорьях. Сюда же могут относиться виды, живущие в условиях холодного, плохо прогревающего даже летом сфагнового болота, их иногда выделяют в отдельную группу оксилофитов.
Формы растений психрофитов многообразны и зависят как от природы организмов, их наследственности, так и от разностороннего воздействия окружающей среды.
Психрофиты можно разделить на 4 группы:
● вечнозеленые деревья и кустарники северных лесов;
● вечнозеленые кустарники тундры и сфагновых болот;
● листопадные кустарнички тундры;
● высокогорные растения и растения каменистых тундр и степей.
К вечнозеленым растениям северных лесов относятся все наши хвойные деревья и кустарники: ель (Picea), сосна (Pinus), сибирский кедр (Pinus sibirica), пихта (Abies), кедровый стланик (Pinus pumila), можжевельник (Juniperus). Все они имеют сильно уменьшенную листовую поверхность. Листья их превращены в иглы (хвою). Структура игольчатого листа типично ксероморфная: это можно объяснить тем, что, сохраняя зимой зеленые листья, хвойные деревья продолжают испарять воду, но лишены возможности быстро насасывать ее из холодной почвы и не могут быстро проводить по древесине, лишенной сосудов. Игольчатые листья являются, таким образом, приспособлением к уменьшению транспирации хвойных растений зимой и ранней весной.
Развитие ксероморфных структур наших хвойных растений, вероятно, определяется произрастанием их или на сухой почве, или на влажных, но холодных субстратах. Всасывание воды здесь ограничивается не только низкой температурой почвы, но и не достаточной ее аэрацией, что ограничивает деятельность корней. В критические периоды временного, но регулярно повторяющегося недостатка воды, растениям полезен ряд выработавшихся у них приспособлений ксероморфного строения.
Вечнозеленые кустарники – обитатели тундр и сфагновых болот. Растения тундры растут в очень суровых условиях. Наличие в почве "вечной" мерзлоты, т.е. слоя, не оттаивающего даже летом, постоянные сильные иссушающие ветры, очень короткий вегетационный период накладывают особый отпечаток на облик растений тундры. Большей частью это приземистые или стелющиеся кустарники с мелкими, обычно вечнозелеными, кожистыми листьями. Мелкие листья, низкорослость – все это результат слабого минерального питания и низких температур, которые задерживают рост растений. Среди многолетних кустарничковых растений тундры встречаются так называемые подушковидные жизненные формы, прижатые к субстрату. Они характеризуются очень слабым годичным приростом своих побегов; в связи с этим старые одревесневшие части побегов прижаты к земле и сильно разветвлены. Поверхность такой низкой подушки сплошь покрываются листьями, которые возникают на многочисленных молодых укороченных побегов. Слабый, медленный рост растений связан и со слабой аэрацией почвы (Богданова Г.А.,1978).
Вечнозеленость многих кустарничков тундры следует рассматривать как полезное свойство, позволяющее растению использовать весной при первых благоприятных условиях перезимовавшие листья для процесса фотосинтеза до появления новых. Ведь вегетационный период очень короток и каждый день важен для накопления необходимых запасов. Таким образом, вечнозеленость здесь приспособление к удлинению вегетации.
Из вечнозеленых тундровых психрофитов можно назвать такие кустарнички:
- луазелеурия лежачая (Loiseleria procumbens);
- кассиопея четырехгранная (Cassiope tetragona);
- вядянка черная (Empetrum nigrum);
- багульник болотный (Ledum palustre).
Кроме вечнозеленности, общим для них является особое строение листьев: они мелкие и завернуты краями вниз так, что прикрывается нижняя поверхность листа с устьицами. Такие листья получили название эрикоидных (от Erica – вереск, у которого листья такого же типа).
Близки к ним по экологии и вечнозеленые кустарнички брусничного типа, с более широкими и толстыми листьями:
- брусника (Vaccinum vitis-idaea);
- подбел (Andromeda polifolia);
- болотный мирт (Chamaedaphne calyculata);
- клюква болотная (Oxicoccus palustris),
произрастающие и на холодных сфагновых болотах.
Во внутренних строениях листьев этой группы психрофитов много черт ксероморфности. Мелкие листья с блестящей толстой кутикулой, часто с густыми волосками на нижней стороне листа, свернутость листовых пластинок – все это давало многим ботаникам повод говорить об этих растениях как ксерофитах. Их относили к особой группе вечнозеленых ксерофитов, приспособленных к "физиологически сухим" почвам, добывание воды из которых затруднено из-за низкой температуры (к таким местообитаниям относятся и сфагновые болота). Интересные опыты, проделанные М.В. Синяниновой-Корчагиной, по выращиванию растений сфагновых болот в лабораторных условиях при достаточном увлажнении при слабом освещении. Оказалось, что багульник, болотный мирт, клюква, подбел чувствовали себя вполне хорошо, а обычно свернутые их листья расправились в широкие пластинки. Прирост побегов у подбела достиг совершенно невстречаемой в естественных условиях высоты – около 0,5 метров.
Исследования В.К. Василевской по онтогенезу листьев психрофитов показывают, что листья их растут и дифференцируют палисадную ткань медленно, за то рано формируют рыхлую губчатую ткань с межклетниками. Такой же тип образования тканей листа наблюдается у листьев субтропических и тропических вечнозеленых растений типа магнолии, т.е. у типичных мезафитов.
Все эти данные не позволяют считать вечнозеленые кустарнички тундры ксерофитами. Можно предполагать, что они произошли от существовавших здесь в третичный период субтропических древесных вечнозеленых растений, т.е. от типичных мезофитов. Суровые условия существования в послеледниковый период сильно изменили облик этих вечнозеленых растений, превратив их в мелколистные стелющиеся кустарнички, которые, однако, вполне приспособились к новым тяжелым условиям обитания.
Третья группа психрофитов тундры – листопадные низкорослые кустарнички:
- дриада точечная (Dryas punctata);
- толокнянка обыкновенная (Arctoctaphylos uvarurs);
- карликовая березка (Betula nana);
- голубика (Vaccinium uliginosum);
- черника (V. Myrtillus).
Это типичные мезофиты, карликовый же рост их связан с низкой температурой и короткой вегетацией.
Строение листьев этой группы растений довольно разнообразно, но степень развития жилок и число устьиц сходны с показателями, характерными для листьев мезофитов.
Среди листопадных кустарничков в тундре встречается много видов многолетних приземистых травянистых растений. Они обладают розеточной или полурозеточной структурой своих побегов.
Наряду с утолщенной стержневой корневой системой дриада точечная (Dryas punctata), лескварелла арктическая (Lesquarella arctica) психрофиты могут быть обладателями хорошо развитой мочковатой корневой системы – злаки, мятлик сизый (Poa glauca), тонконог азиатский (Koeleria asiatica) – или могут развивать толстые корневища с придаточными корнями.
Цветки у психрофитных растений семейства вересковые самой разнообразной величины, обычно собраны в кистевидное соцветие на концах ветвей, реже соцветие бывает щитковидное (у багульника или рододендрона). Большинство вересковых имеет 5 чашелистиков, 5 лепестков, 10 тычинок, в завязи 5 гнезд, на рыльце можно рассмотреть 5 лучей. Однако четырех членный план цветка считается вполне типичным для семейства (у вереска, брусники, клюквы, дабеции (Dabecia)). Встречаются также двучленные (у симпиезы (Sympieza)) и трехчленные цветки (у кладотамнуса (Cladothamnus)), а у бежарии (Bejaria) они, как правило, семичленные. Хотя почти у всех вересковых цветки актиноморфные, в отдельных случаях наблюдаются уклонения от лучевой симметрии. У некоторых видов рододендрона, например, при 5 чашелистиках число лепестков 3 и они образуют зев. У некоторых вересковых (например, у багульника) лепестки свободные. Однако у большинства видов лепестки срастаются при основании или до половины своей длины так, что, потянув за один лепесток, мы отделим весь венчик. Наличие нектарного диска является характерным признаком семейства, хотя и в данном случае не обходится без исключений. У некоторых видов эрики диска нет. Обычно тычинки расположены двумя кругами: один – против лепестков, другой – против чашелистиков. Общее для всех тычинок только то, что они двугнездны и вскрываются чаще всего порами. (Приложение 1, рис.1).
Пыльцевые зерна расположены в тетраэдрических тетрадах, имеют почти шаровидную форму. Длина полярной оси отдельных пыльцевых зерен 23-30мкм, экваториальный диаметр 30-39мкм. В очертании тетрады округло-треугольные, 42-45мкм в диаметре. Борозды соприкасаются между собой, длина полуборозды 13-17мкм, ширина 4,5-6,5мкм; края борозд ровные, концы слегка оттянуты, притуплены и направлены в сторону дистальной поверхности; мембрана борозд гладкая. Экзина покровная, толщиной 0,7-1,0мкм. Пыльца темно-серого цвета (Приложение 1, рис.2).
В виде тетрад пыльца и попадает на рыльце, где все пыльцевые зерна сразу прорастают. Тетрады образуются у насекомоопыляемых растений, тогда как ветроопыляемые вересковые имеют пыльцу из одиночных зерен.
Весьма характерным типом цветка для вересковых является бокальчатый цветок. Он присущ как ягодным вересковым (бруснике, чернике), так и неягодным, которых большинство. У клюквы, однако, полностью развившийся цветок напоминает цикламен, так как лепестки у него заворачиваются назад, а вперед выставлен длинный гинецей, к столбику которого тесно прилегают длинные пыльники; цветки при этом поникающие. О.Хагеруп изолировал соцветия многих вересковых с помощью мешочков и убедился, что, не смотря на невозможность перекрестного опыления, плоды у них образовывались, но при условии, что цветки встряхивались. В природе встряхивание производит ветер, и по этому ягод черники или голубики меньше в тех местах, где деревья сближены и гасят ветер. Плоды вересковых представляют собой септипцидную или локулицидную коробочку, ягоду или же костянку (Боч М, С., Смагин В, А., 1993).
Семейство вересковые (Ericaceae): их классификация и морфологическая характеристика.
Семейство вересковые очень не однородно и делится на ряд подсемейств. В 1971 г. Английский ботаник П.Ф. Стивене предложил новую систему этого семейства, которая имеет ряд преимуществ по сравнению со старыми системами. В его системе вересковые делятся на 6 подсемейств.
Наиболее примитивно подсемейство рододендровых (Rhododendroideae), куда входят исключительно деревья и кустарники. Венчик у них свободнолепестный или сростнолепестный, обычно с широким зевом. Тычинки без придатков; пыльники вскрываются верхушечными порами или короткими, или длинными щелями. В пыльниках обычно имеются шелковистые нити, связывающие пыльцу в комочки. Завязь 2 – 14- гнездная, часто покрытая железистыми волосками. Плоды – коробочки, обычно септицидные. Семена обычно многочисленные. В подсемейство входят американский род бежария (около 30 видов), рододендрон (вероятно, до 800 видов), мензисия (7 видов), багульник (около 10 видов в северной умеренной зоне и в Арктике), эпигея (3 вида), кальмия (8 видов), филлодоце (7 видов) родотамнус (2 вида), монотипные роды бриаитус и луазелерия и некоторые другие. Наиболее известен самый крупный в подсемействе род – рододендрон, приобретший исключительное значение в декоративном садоводстве. Отдельными островками он встречается на западе европейской части России. Из дальневосточных рододендронов особый интерес представляют рододендрон Фори (R. fauriei) и рододендрон Шлип-пенбаха (R. schlippenbachii) (Сабинин Д, А., 1963).
В подсемейство эриковые (Ericoideae) входят вечнозеленые кустарники с мелкими листьями с сильно завернутыми краями. Венчик всегда сростнолепестный, остающийся. Пыльники вскрываются очень крупными порами или щелями; придатки пыльников, когда они имеются, представляют собой шпоры; шелковистые нити в пыльниках отсутствуют. Плод обычно – локулицидная коробочка. Семян в каждом гнезде от одного до многих. Сюда относятся, прежде всего, большой род эрика (более 500 видов), монотипный род вереск и другие.
Обширное подсемейство вакциниевые (Vaccinioideae) включает древесные растения очень разного облика – от небольших деревьев до маленьких кустарничков. Венчик обычно сростнолепестный, редко свободнолепестный, обычно кувшинчатый или трубчатый. Пыльники очень разных типов, лишены шелковистых нитей. Завязь верхняя или нижняя, 3-10-гнездная. Плод – ягода, костянка или локулицидная коробочка, обычно с несколькими семенами в гнезде, редко с одним. Сюда относятся арбутус (Arbutus, 20 видов), Кассиопея (12 видов), энкиантус (Enkianthus, 10 видов), подбел (1-2 вида в умеренных и холодных областях северного полушария), арктерика (Arcterica, 1 вид), гаультерия (Gaultheria, около 200 видов), вакциниум (Vaccinium, около 350 видов, клюква встречается многими авторами в вакциниум) и др. далеко не всеми признаваемый и часто объединяемый с вакциниумом род клюква (Oxycoccus) содержит не более 4 видов, распространенных в умеренных и холодных областях северного полушария. Вечнозеленые, большей частью стелющиеся мелкие кустарники с тонкими стеблями, мелкими кожистыми листьями и темно-красными ягодами.
Подсемейство грушанковые (Pyroideae) небольшое; в него входят 4 рода и около 30 видов, представленные в умеренных и холодных областях северного полушария. Можно даже сказать, что это в основном таежное подсемейство, т.к. именно в таежной зоне сосредоточена большая часть его представителей. В отличие от всех ранее рассмотренных деревянистых вересковых грушанковые – травянистые вечнозеленые растения. У большинства видов имеется прикорневая розетка глянцевитых листьев, из которых выходит цветочная стрелка с кистевидным соцветием. Цветки 4-5-членные, открытые или бокальчатые. Последние имеют прямой столбик; у открытых он длинный, изогнутый, противопоставленный 10 (реже 8) тычинкам. Плод у грушанковых – сухая коробочка с большим количеством очень мелких семян, лишенных семядолей, и с зародышем, который состоит из нескольких клеток. Микориза для существования грушанковых еще более необходима, чем для других вересковых. Существует предположение, что продукты фотосинтеза необходимы грушанковым только для образования семян, а другие нужды растения вполне удовлетворяются питанием с помощью микоризы. Все представители грушанковых имеют длинное корневище и только у одноцветки оно заменено многолетними корнями.
К подсемейству вертляницевых (Monotropoideae) относятся 10 родов и 12 видов. Большинство родов этого подсемейства монотипны. Вертляницевые полностью перешли на питание с помощью микоризы, в связи, с чем утратили зеленую окраску. Многие из них имеют тусклый землистый цвет, но некоторые окрашены ярко. Так, монотипный род саркодес (Sarcodea), произрастающий на юго-западе США, где его называют снежным цветком, имеет малиновую окраску всего растения. Вертляницевые имеют толстый мясистый стебель с листьями в форме чешуи. Цветки у вертляницевых собраны в конечное поникающее соцветие, за исключением подъельника одноцветкового (M.uniflora). цветки 5-членные или реже4-8-членые. Цветок имеет колокольчатую форму, но у одних представителей лепестки срастаются у основания, а у других остаются свободными. Нектарный диск имеется у большинства представителей этого подсемейства. Плод – коробочка, часто перепончатая. Мелкике семена птероспоры (Pterospora) снабжены крылышком, значительно превосходящим по размеру само семя.
Подсемейство виттштейниевых (Wittsteinioideae) с монотипным австралийским родом виттштейния (Wittsteinia) связывает вересковые с семейством эпакрисовые. Виттштейния – это мелкий вечнозеленый кустарник со стелющимися побегами и мясистым плодом.
2.3 Обзор видового состава вересковых (Ericaceae) Красноселькупского района
Багульник (Ledum). Русское название "багульник" происходит от старинного глагола "багулить", что означает "отравлять". В русском названии отображена характерная особенность этого кустарника – сильный, удушающий запах. Научное же название "ледум" (Ledum) происходит от греческого ledon - так древние греки называли растение, из которого добывалась ароматическая смола – ладан (Ladanum)
Багульник болотный – Ledum palustre. Индицирует бедность условий местопроизрастания. Широко распространен в природе и чаще других встречается в культуре. Растет в тундре и лесотундре, на торфяниках, на верховых болотах, в подлеске сырых хвойных лесов, вдоль горных речек и ручьев, в высокогорьях, также среди кедрового стланика небольшими группами, зарослями.
Сильноветвистый вечнозеленый кустарник высотой от 50 до 120см, с приподнимающимися побегами, покрытыми густым "ржавым" войлочным опушением. Диаметр куста во взрослом состоянии около 1 метра. Листья ланцетные, темные, блестящие, с запахом. Края листьев сильно повернуты вниз. Цветки (до 1,5см в диаметре) белые, реже розоватые, остропахнущие, в многоцветковых зонтиках (июнь-июль). Плод – коробочка раскрывается пятью створками. Семена созревают в середине августа. Корни поверхностные, с микоризой. Все части растения ядовиты! Во время цветения выделяют в воздух вещества, которые в больших количествах оказывают неблагоприятное воздействие на человека, выражаясь сильной головной болью.
На моховых болотах багульнику грозит опасность быть затянутым в толщу мха, т.к. мох ежегодно нарастает и поверхность болота неизменно повышается. Таким образом, корень и большая часть побегов со временем оказываются в переувлажненном, лишенном кислорода слое и постепенно отмирают, перегнивают и становятся частью торфа, а на живых побегах, над поверхностью мхов, образуются придаточные корни.
Багульник гренландский – Ledum decumbens. Растет в кустарничковых тундрах на кочкарниках с редколесьем, на песчаных холмах, в зарослях кедрового стланика, на высокогорных сфагновых болотах, каменистых россыпях. Вечнозеленый кустарничек 20-30см высотой. Цветет необильно, но ежегодно со второй декады мая до середины июня. Плодоносит нерегулярно. Семена созревают в конце августа. Растет медленно, ежегодный прирост около 1см.
Багульник крупнолистный – Ledum macmphyllum. Растет в подлеске хвойных лесов, на сфагновых болотах, по окраинам каменных россыпей среди зарослей вересковых кустарничков. Вечнозеленый кустарник до 1,3м высотой. Цветет обильно со второй половины мая до первой декады июня. Семена созревают в конце августа – начале сентября. Ежегодный прирост 3-4см, редко 6-8-см.
Брусника (Vaccinium). Крупный род, включающий такие растения как клюква, брусника, черника и голубика. Все они предпочитают кислую почву и образуют прямостоячие или стелющиеся кустики. В заболоченные сырые и влажные пойменные и террасовые местоположения заходит редко и единичными экземплярами. Существуют и карликовые формы, и растения высотой около 1м и более.
Брусника обыкновенная (Vaccinium vitis-idaea L.) – длиннокорневищный вечнозеленый кустарничек из семейства вересковых. Индицирует относительно бедные и свежие условия.
Удлиненные побеги брусники полициклические и приподнимающиеся. Листья кожистые, многолетние, очередные, эллиптические или обратнояйцевидные, тупые или слегка выемчатые, слегка зазубренные или цельнокрайние с завороченным краем 5-27мм длиной, сверху темно-зеленые, снизу бледные с темно-бурыми рассеянными железками.
Основание побега переходит в гипогеогенное корневище, залегающее на глубине 2-10см, обычно очень длинное – до 18м длиной, шнуровидное, зеленовато-коричневого цвета. От корневища отходят тонкие короткие и редко расположенные придаточные корни.
Соцветие – короткая густая поникающая кисть с 2-8- (редко 18) цветками, формируется в верхней части прошлогоднего годичного побега. Цветки на коротких опушенных красноватых цветоножках, которые обладают слабым, но приятным запахом. Чашечка спайнолистная четырехзубчатая, с короткими округлыми красноватыми зубцами. Венчик спайнолепестный колокольчатый, бледно-розовый с четырьмя лопастями. Тычинок – 8 с волосистыми тычиночными нитями, пыльники без придатков. Столбик выдается из венчика, завязь четырехгнездная, гинецей синкарпный из четырех плодолистиков, плацентация центрально-угловая.(Баландина, Муратов, 1978).
Брусника имеет обширный голарктический ареал с преимущественным распространением в северной части Евразии. Встречается по всей территории СНГ, кроме южных районов европейской части, всей Средней Азии, подавляющей части Казахстана и Закавказья (Атлас ареалов..., 1976).
Брусника растет в лесной и арктической зонах, поднимается до гольцового пояса, распространена в хвойных и смешанных лесах, на вырубках, в горных и равнинных тундрах. Чаще всего она встречается в травяно-кустарничковом ярусе сосновых и лиственничных лесов, являясь доминантом этого яруса (Брусника.., 1986).
Голубика (Vaccinium). Иногда голубику называют пьяника, или гонобобель за то, что она якобы пьянит и гонит боль в голову. На самом деле виновник этих явлений багульник, часто растущий по соседству с голубикой.
Крупный род, включающий такие растения как клюква, брусника, черника и голубика. Все они предпочитают кислую почву и образуют прямостоячие или стелющиеся кустики.
Голубика обыкновенная, топяная, гонобобель – Vaccinium uliginosum L.голубика в естественных условиях произрастает в лесах, заболоченных или каменистых тундрах, на бедных кислых почвах, кочках болот. Внутри вида различают несколько подвидов, каждый из которых произрастает в своем эколого-географическом районе.
Многолетний листопадный, сильноветвистый кустарник высотой до1м, с прямостоячими цилиндрическими ветвями, с буроватой или темно-серой корой, побеги зеленые. Листья очередные, гладкие, жесткие, мелкие до 3см длиной и до2-4 см шириной на очень коротких черешках, от обратнояйцевидных до ланцетных, на верхушке тупые, с цельными или немного загнутыми вниз краями, с негустыми стебельчатыми железками, сверху голубовато-зеленые, покрыты восковым налетом, снизу более светлые и с сильно выступающими жилками. Цветки поникающие, кувшинчато-колокольчатые длиной до 6см, сидящие по 2-3- на верхушках прошлогодних веточек. Цветоножки обычно несколько длиннее цветка, с двумя неравными длиной 2-5 мм, пленчатыми, зеленоватыми прицветниками. Чашечка состоит из 4-5 округлых маленьких чашелистиков. Венчик кувшинчатый, беловатый с отгибом из коротких отогнутых наружу зубцов. Тычинок 8 или 10, завязь 4-5-гнездная, столбик длиннее тычинок. Ягода разнообразная по форме, чаще продолговатая, длиной 1,2см, синяя с сизым налетом, массой до 0,8г. Семена многочисленные, длиной до 1,5мм, светло-коричневые, полулунной формы. Цветет в июне, продолжительность цветения 10-12 дней, ягоды созревают через 50 дней после зацветания (Крылов А.Г., 1984).
Клюква (Oxycoccus). В мире 4 вида клюквы произрастают в тундре, лесотундре и в лесной зоне Европы, Азии и Северной Америки. В России в естественных условиях произрастают 2 вида (болотная и мелкоплодная). Произрастает клюква на верховых болотах в виде небольших стелющихся укореняющихся кустиков на почти лежачих цветоносах, на которых и образуются крупные красные ягоды, кажущиеся прямо лежачими на сфагновом мху.
Клюква четырехлепестная, обыкновенная, болотная – Oxycoccus macrocarpus.
Растет на сфагновых и осоково-сфагновых болотах, в тундре, лесотундре и лесной полосе европейской части страны, в Сибири, на Дальнем Востоке, на Камчатке и острове Сахалин.
Вечнозеленый стелющийся ветвистый кустарничек, со стелющимися длинными тонкими и гибкими одревесневающими темно-бурыми стеблями, длиной до 75см и около 1мм в диаметре, с приподнимающимися цветоносными веточками и короткими нитевидными пушистыми однолетними веточками. Листья на коротких черешках, кожистые, продолговато-яйцевидные, до 1,6 см длиной, до 0,6 см шириной, на верхушке острые с цельно завороченными краями, сверху темно-зеленые и блестящие, снизу сизоватые от воскового налета с выдающейся снизу срединной жилкой. Цветки поникающие, расположенные по 1 или по2-4 в зонтиковидных соцветиях на прошлогодних ветвях. Цветоножки длинные (до 4,5см), одноцветковые, выходящие из пазух чешуевидных верхушечных листочков, красноватые, пушистые, несущие по 2 маленьких острых прицветника длиной около 1мм. Чашечка с 4 округлыми, по краям тонко- и длиннореснитчатыми чашелистиками длинной около 1мм, сначала зеленая, затем - красная. Венчик глубокочетырехраздельный, длиной 5-7мм и шириной 1,5-2мм, с розово-красными, загнутыми наружу назад продолговато-ланцетными долями длиной 4-6мм. Тычинок 9, с короткими нитями почти равными им пыльниками. Завязь четырехгнездная, столбик немного длиннее тычинок и вместе с ними выделяется из венчика. Ягода шаровидная, крупная, 0,8-1,8см в диаметре. Масса одной ягоды 0,5-1,5г, темно-красная, сочная, кислая, четырехгнездная, многосемянная, на плодоножке, перезимовывающая. Семена длиной 1,5-2,8мм, яйцевидной формы. Цветение – с конца мая по начало июня. Ягоды созревают (у ранних и среднеспелых сортов) в конце августа – начале сентября, т.е. на 2-3 недели раньше клюквы крупноплодной. Однако размер приростов и сила роста кустов клюквы болотной значительно (почти вдвое) меньше. Весной, после схода снега можно собирать оставшуюся зимовать так называемую подснежную клюкву, более сладкую, но менее транспортабельную и хуже хранимую. Обильное плодоношение периодично. Этот вид характеризуется большой внутривидовой изменчивостью растений по форме и величине листа, числу цветков в соцветии, величине, форме и окраске плодов.
Клюква мелкоплодная – Oxicoccus microcarpus.
Населяет тундровую и лесную зоны, горнотунлровый, альпийский и субальпийский пояса гор северного полушария. Клюква мелкоплодная очень похожа на клюкву четырехлепестную, но отличается от нее более мелкими размерами. Многолетний вечнозеленый стелющийся полукустарник с тонкими побегами длиной до 50см и толщиной до 0,6мм. Листья длиной до 0,7см и шириной до 0,2см, яйцевидные, заостренные, с сильно завернутыми краями. Цветки розовые или темно-розовые, похожи на крошечные цветы цикламена. Ягоды шаровидные или продолговато-яйцевидные, 3-10мм в диаметре, масса ягоды 0,2-0,3г. В соцветии формируется до 3 ягод. Цветет в июне, июле. Плодоносит в сентябре (Работнов Т.А., 1978).
Глава III. Морфо-анатомические и экологические особенности строения психрофитных растений семейства вересковые
Вересковые широко распространены по земному шару, большинство представителей вересковых – кустарники или кустарнички, иногда травы, но среди них имеется и крупные деревья. Кустарники и кустарнички из вересковых встречаются и стелющейся формы, такова всем известная клюква (Oxycoccus). Некоторые виды маклинии (Mackleania) и агапетеса (Agapetes) представляют собой кустарники с длинными цепляющимися побегами. Среди азиатских видов рододендрона, а также среди вакциниевых, в том числе маклинии, известны эпифиты. К ним относятся рододендрон камелиецветный (R.camelliiflorum), рододендрон голубичный (R.vaccinioides), рододендрон деревцовый (R.dendricola) и др. под верхним слоем эпидермы листья эпифитов имеют слой крупных тонкостенных клеток, служащих для запасания воды. Этот признак отсутствует у вересковых, ведущих наземный образ жизни. Семена у некоторых видов снабжены своеобразным придатком, похожим на летучки, что является приспособлением в данном случае не для переноса, а для зацепки за ветви деревьев, на которых они начинают развиваться. Мы в своей работе остановимся на описании тех представителей семейства вересковые, которые относятся к группе психрофитных растений и обитают в тайге и тундре (Юдина В.Ф., Белоногова Т.В., Колупаева С.Н., 1986).
Облик вересковых весьма своеобразен, поэтому в ботанике существует понятие "эрикоидный облик". Эрикоидный облик создают деревянистые побеги, покрытые мелкими кожистыми листьями. Во многих случаях листья концентрируются на концах побегов, что хорошо видно у рододендронов. У одних видов листья совершенно голые и имеют глянцевитую поверхность, у других они по-разному опущены. Опущение развивается обычно на нижней поверхности листа, где у большинства вересковых находятся устьица (Мазная Е.А., 2001).
Потянув за кустик многих кустарничков из семейства вересковых – черники оыбкновенной (Vaccinium myrtillus), подбела (Andromeda), брусники (Vaccinium vitis-idaea), хамедафне болотной (Chamaedaphnecalyculata), мы обычно видим, что от кустика отходит длинное корневище в виде шнура, которое вскоре подводит нас к следующему кустику, а далее – к следующему, и оказывается, что большая группа кустиков имеет общее корневище. У взрослой черники обыкновенной оно достигает 3 метра. У других черник корневище тянется на 10 метров.
У многих вересковых (кустарничков и низких кустарников) в почве скрыт почти полностью стволик кустика, например у багульника (Ledum). У третьих укореняются полегающие ветви, например у кассиопеи (Cassiope), вереска (Calluna).
Древесина у вересковых очень плотная и часто смолистая, напоминающая древесину хвойных. Смолы содержатся и в листьях, иногда выступая на поверхность.
3.1 Материалы и методы исследования
При изучении анатомического строения стеблей и листьев психрофитных растений семейства вересковые нами был изготовлен ряд временных микропрепаратов. Для этого изготавливались поперечные срезы из свежих стеблей багульника болотного, голубики обыкновенной и болотного мирта. Срезы делали зажав стебли в сердцевине бузины. Отобрали наиболее тонкие, и на одни из них действовали хлор-цинк-йодом, на другие – флорглюцином и соляной кислотой. Затем, оттянув реактивы фильтровальной бумагой, клали срезы в каплю глицерина. При изучении анатомического строения листьев срезы делали перпендикулярно главной жилке. Срезы делали, зажав объекты в сердцевине стебля бузины. Помещали их в каплю воды и при малом увеличении отбирали наиболее тонкие. Затем проводили реакцию на лигнин, клали в каплю глицерин на предметное стекло, накрывали покровным и приступали к изучению.
После изучения микроскопического строения объектов зарисовали их. При изготовлении рисунков сохраняли пропорции, отмечая взаиморасположение и размеры тканей и клеток их слагающих.
3.2 Анатомические особенности строения вечнозеленых и листопадных психрофитных кустарничков семейства вересковые
Общие анатомические признаки у вересковых проявляются, прежде всего, в строении стебля. Так нами изучались строения стебля голубики, багульника и болотного мирта. Все они отличаются общими особенностями в строение стебля, которые заключаются, прежде всего, в следующем:
● стебель имеет четко выраженные три блока – первичная кора, центральный цилиндр и сердцевина;
● первичная кора образована толстостенными клетками колленхимы, где располагаются хорошо выраженные крупные воздухоносные полости;
● флоэма и ксилема представлены сплошными кольцами, т.е. стебель имеет типичное непучковое строение с кольцевым способом заложения камбия;
● сердцевина достаточно крупная, образована паренхимными клетками.
Снаружи стебель багульника покрыт однослойной покровной тканью – эпидермой – клетки ее имеют несколько вытянутую форму, плотно сомкнуты между собой. Далее располагается механическая ткань (предположительно уголковая колленхима) с сильно утолщенными клеточными стенками и малыми внутренними полостями. Между клетками первичной коры имеются достаточно крупные воздухоносные полости. Центральный цилиндр представлен многослойной флоэмой и трехслойной, в нашем случае исследовался трехлетний побег, ксилемой. Ксилема образована толстостенными прозенхимными клетками с достаточно сильно утолщенными клеточными стенками. В поперечном сечении форма клеток близка к прямоугольнику. Между клетками ксилемы и флоэмы располагается камбий, образующий вторичные проводящие ткани. Центральное положение занимает сердцевина, имеющая округлую форму и состоящая из очень крупных паренхимных клеток с неутолщенными клеточными стенками. Ее клетки выполняют запасающую функцию (рис.1.).
Стебель голубики имеет свои анатомические особенности, отличающие его от выше описанного стебля багульника (рис.2).
Эпидерма так же однослойная, но клетки более мелкие, чем у стебля багульника. Далее закладывается механическая ткань, уголковая колленхима с утолщенными клеточными оболочками и более крупными внутренними полостями, по сравнению с клетками стебля багульника. Воздухоносные полости примыкают непосредственно к флоэме, в отличие от стебля багульника, где они были полностью окружены механической тканью.
Флоэма состоит не только из ситовидных трубок, но включает в себя и лубяные волокна. В ксилеме хорошо выражены сердцевинные лучи, идущие от флоэмы к сердцевине стебля. Сердцевина имеет форму пятиугольника и образована клетками правильной паренхимной формы, имеющими небольшие межклетники.
Стебель болотного мирта снаружи покрыт однослойной эпидермой, клетки колленхимы утолщены незначительно, соответственно увеличиваются их внутренние полости. Воздухоносные полости в первичной коре самые крупные из всех изученных нами стеблей, близки по форме к прямоугольникам. Флоэма слабо развита, ситовидные трубки почти не просматриваются, в основном, она состоит из клеток лубяных волокон. Зато слой ксилемы развит достаточно мощно, хорошо выражены сердцевинные лучи, паренхима сердцевины одревесневшая, образована крупными плотносомкнутыми клетками (рис.3).
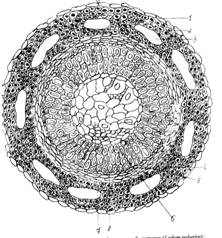
Рис.1. Анатомическое строение стебля багульника болотного (Ledum palustre): 1 – эпидерма; 2 – уголковая колленхима; 3 – воздухоносные полости; 4 – флоэма; 5 – вторичная ксилема; 6 – первичная ксилема; 7 – сердцевина; 8 – камбий.
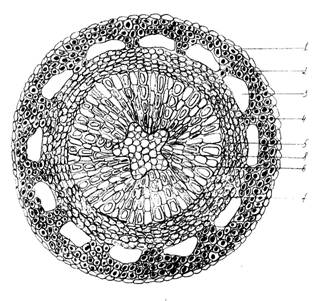
Рис.2. Анатомическое строение стебля голубики обыкновенной (Vaccinium uliginosum L.): 1 – эпидерма; 2 – колленхима; 3 – воздухоносные полости; 4 – флоэма; 5 – камбий, 6 – сосуды ксилемы; 7 – сердцевинные пучки; 8 – сердцевина.
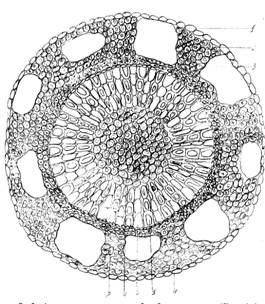
Рис. 3. Анатомическое строение стебля болотного мирта (Chamaedaphne calyculata): 1 – эпидерма; 2 – колленхима; 3 – воздухоносные полости; 4 –флоэма; 5 – ксилема; 6 – сердцевинный луч; 7 – сердцевина; 8 – камбий.
В общих чертах анатомическое строение стеблей вересковых соответствует условиям их произрастания – переувлажненным местам обитания с холодными почвами, что способствует развитию воздухоносных полстей. Поэтому они имеют признаки ксероморфной структуры. Это проявляется и в сплошном заложении проводящей ткани и в развитии механической ткани.
В анатомическом строении листьев растений семейства вересковых так как и стебля имеется значительное разнообразие, так встречаются вечнозеленые и листопадные виды вересковых.
Среди разнообразия листьев вересковых выделяют 3 типа:
● тип рододендрона – лист плоский или со слегка завернутыми вниз краями.
Такой лист имеют клюква, брусника, голубика и др.;
● игловидный лист – напоминает хвоинку как у некоторых видов эрики (Erica),
хариманеллы (Harrimanella);
● эрикоидный лист с сильно завернутыми книзу краями (у вереска).
У некоторых вересковых листья вообще уникальны. Так, кассиопея Редовского (C.redowskii) имеет лист, напоминающий палец перчатки, т.е. с почти замкнутой полостью. У многих вересковых лист может свертываться при наступлении засушливого периода; это отмечено, в частности, для брусники (Тимошок Е.Е., Паршина Н.В., 1992).
Таким образом, у ряда вересковых существуют динамические реакции на изменения погодных условий. Чрезмерное испарение воды в сухую погоду ослабляет напряжение в слоях мезофилла на нижней стороне листа, поэтому лист выгибается, скручивается; контакт листовой полости с окружающей средой уменьшается. Имеется, однако, сведения, что у очень многих вересковых, в частности у самого вереска, устьица есть и на верхней стороне листа. Таким образом, испарение происходит даже при полностью закрытой листовой полости. Значение этой полости становится понятным, если лист опустить в воду.
Анатомическое изучение листьев этих растений обнаруживает, что и "ксероморфность" их далеко не так ярко выражена как кажется.
Нами были изучены строение листьев некоторых психрофитных вечнозеленых и листопадных кустарничков. Так, например, при рассмотрении поперечного среза листа багульника мы обнаружили, что эпидерма однослойная, образована мелкими, плотно сомкнутыми клетками, на внешней стороне, контактирующей с атмосферой, откладывается толстый слой кутикулы. На нижней поверхности листа расположено большое количество простых и головчатых железистых волосков, устьица здесь не углубленные, а поверхностные. Число устьиц на единицу поверхности невелико.
Мезофилл листа гомогенный, имеет сильно развитую губчатую ткань, среди клеток которой находится большое количество межклетников. Сеть жилок не густая, проводящие ткани пучков развиты не сильно. Механических тканей, столь характерных дл листьев ксерофитов, почти нет (рис.4).
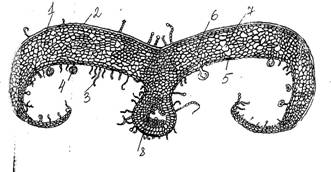
Рис.4 Поперечный срез листа багульника болотного (Ledum palustre): 1 – эпидерма; 2 – кутикула; 3 – кроющие волоски; 4 – железистые волоски; 5 – устьице; 6 – губчатый мезофилл; 7 – межклетники; 8 – закрытый коллатеральный проводящий пучок.
Также нами изучено строение листьев клюквы, относящейся к группе вечнозеленых кустарничков и голубики, принадлежащей к листопадным кустарничкам. В результате анатомических исследований мы выяснили, что анатомическое строение данных листьев имеет достаточное количество общих черт:
● Эпидерма однослойная, на внешней стороне верхней эпидермы имеет толстый защитный слой кутикулы;
● Устьица располагаются только на нижней стороне листа, среди клеток нижней эпидермы и на одном уровне с ее клетками, т.е. не являются погруженными;
● Мезофилл гетерогенный, четко дифференцирован на хорошо развитую палисадную ткань, как правило, двухслойную и губчатую;
● В губчатом мезофилле достаточное количество крупных межклетников и воздухоносных полостей, примыкающих к устьичным аппаратам.
Эпидерма четко подразделяется на нижний слой, состоящий из более мелких клеток, среди которых находятся устьица и верхний образованный крупными плотносомкнутыми клетками. Отличия же заключаются в том, что клетки эпидермы у листа голубики мельче, чем у листа клюквы, у которой отмечен и достаточно мощный слой кутикулы. Клетки мезофилла наоборот крупнее у листа голубики, палисадный мезофилл состоит из вытянутых клеток расположенных в два слоя, клетки губчатой ткани также имеют вытянутую форму, образуют межклетники и располагаются перпендикулярно нижней эпидерме (рис.5).
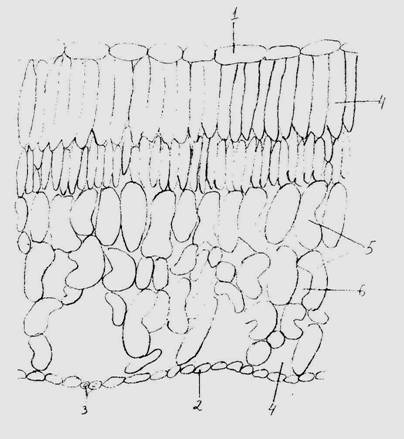
Рис.5. Поперечный срез листа голубики обыкновенной (V. Uliginosum L.): 1 – верхняя эпидерма; 2 – нижняя эпидерма; 3 – устьица; 4 – палисадный мезофилл; 5 – губчатый мезофилл; 6 – межклетники; 7 – воздухоносные полости.
Далее в своей работе мы изготовили поперечные срезы с листьев однолетнего и прошлогоднего листа клюквы и сравнили их строение (рис.6).
При изучении полученных микропрепаратов мы обнаружили, что в однолетнем листе клетки эпидермы мельче, а слой кутикулы тоньше. У листа второго года жизни эпидермальные клетки, особенно входящие в состав нижней эпидермы, значительно крупнее, а слой кутикулы толще почти в два раза. Мезофилл в первом случае слабо дифференцирован на слои палисадной и губчатой ткани и, в основном, представлен рыхлой губчатой тканью с достаточно крупными межклетниками. Лист второго года жизни имеет гетерогенный мезофилл, четко подразделяющийся на верхний палисадный слой, образованный двумя рядами клеток и нижний губчатый с межклетниками и воздухоносными полостями.
Все особенности строения изученных нами листьев не позволят считать их листьями типичного ксерофита: у них есть признаки мезоморфной и даже гигроморфной структуры. Таким образом, эта группа растений по своему анатомическому строению ближе к мезофитам.
Достаточное водоснабжение, но слабое минеральное питание, плохо аэрируемая и холодная почва вызывают общую задержку их роста, а произрастание на открытых, не защищенных от ветра зимой и хорошо освещаемых летом местах, привело к образованию у них мощной кутикулы, играющей роль экрана, отражающего яркие солнечные лучи и уменьшающего испарение. Интенсивным освещением можно объяснить и сворачивание листьев, которое уменьшает транспирацию.
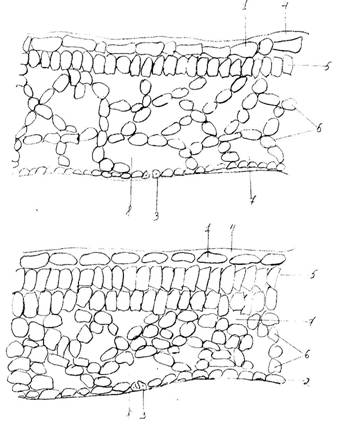
Рис.6. Поперечный срез листа клюквы обыкновенной (Oxycoccus
macrocarpus): 1 – верхняя эпидерма; 2 – нижняя эпидерма; 3 – устьица; 4 – кутикула; 6 – губчатый мезофилл; 7 – межклетник; 8 – воздухоносные полости.
ЗАКЛЮЧЕНИЕ
Специфика природных условий Севера и, в частности, экстремальные значения целого ряда жизненно важных для растений экологических факторов не могли не сказаться (и заметно сказались!) на морфологических, эколого-физиологических и биологических признаках, свойствах и особенностей северных растений. По совокупности данных показателей выделили отдельную экологическую группу растений – это растения психрофиты.
К ним относят виды, произрастающие в тайге, тундре, на высокогорьях. Сюда же могут относиться растения, живущие в условиях холодного, плохо прогревающегося даже летом, сфагнового болота.
Форма растений психрофитов многообразны и зависят как от природы организмов, так и от разностороннего воздействия окружающей среды.
Среди психрофитных растений встречаются представители семейств: осоковые, гвоздичные, крестоцветные, сложноцветные, большое количество видов семейства вересковые. Большинство вересковых – это вечнозеленые и листопадные кустарнички тундры и сфагновых болот.
Растения тундры живут в очень суровых условиях, что накладывает особый отпечаток на их облик. Большей частью – это приземистые или стелющиеся кустарнички с мелкими, обычно вечнозелеными, кожистыми листьями. Кроме вечнозелености, общим для них является особое строение листьев: они мелкие и завернуты краями вниз так, что прикрывается нижняя поверхность листа с устьицами.
Листья, в основном, мелкие, эпидерма однослойная с блестящей толстой кутикулой, часто с густыми волосками на нижней стороне листа. Число устьиц на единицу поверхности невелико. Мезофилл листа, в ряде изученных нами случаев, – гетерогенный, иногда имеет только губчатую ткань с большим количеством межклетников. Сеть жилок негустая, проводящие ткани пучков развиты несильно, механических тканей почти нет.
Общее анатомические признаки у психрофитных растений семейства вересковых проявляются и в строении стебля. Все изученные нами стебли отличались общими особенностями:
● стебель имеет четко выраженные 3 блока: первичная кора, центральный цилиндр и сердцевина;
● первичная кора образована толстостенными клетками колленхимы, где располагаются хорошо выраженные крупные воздухоносные полости;
● флоэма и ксилема представлены сплошными кольцами, т.е. стебель имеет типичное непучковое строение, с кольцевым способом заложения камбия.
Библиография
1. Алексеев С.В., Беккер А.М. Изучаем экологию экспериментально. – СПб.: Феникс, 1993. – 201с.
2. Алексеев Ю.Е., Баландин С.А. Энциклопедия растений России. Растения тундры. – М.: Классикс Стиль, 2003. – 208с.
3. Атлас ареалов и ресурсов лекарственных растений СССР – М.: ГУГК, 1976. – 340с.
4. Бавтуто Г.А., Ерей Л.М. Практикум по анатомии и морфологии растений. – Минск: Новое знание, 2002. – 458 с.
5. Богданова Г.А., МуратовЮ.М., Брусника в лесах Сибири. – Новосибирск: Наука, 1978. – 178с.
6. Большой атлас природы России / Под ред. М.Сергеевой, С.Бровкина. – М.: Эгмонт Россия Лтд, 2003. – 644 с.
7. Боч М.С., Смагин В.А. Флора и ратительность болот Северо-запада России и Принципы их охраны. – СПб.: Гидрометеоиздат, 1993. – 224с.
8. Брусника: Морфология и анатомия. Фитоценотическая приуроченность. Урожайность. Хранение и переработка. Химический состав ягод / Юдина В.Ф., Белоногова Т.В., Колупаева С.Н. и др. – М.: Наука, 1986. – 73с.
9. Бумар Г.И., Таргонский П.Н. Возрастной состав ценопопуляций Vaccinium vitis-idaea L. в Полесском государственном заповеднике (УССР) / Ратительные ресурсы. – СПб.: ГУГК 1989. – Т.25, вып.3. – с.349-353.
10. Викторов В.П., Гуленькова М.А., Дорохина Л.Н. и др. Практикум по анатомии и морфологии растений. – М.: Академия, 2001.
11. В краю селькупов: Красноселькупский район: природа.Люди.Экономика. экология / кол.авторов. – Екатеринбург: Сред.-Урал.кн.изд-во, 2000. -200с.
12. Ипатов В.С., Кирикова Л.А. Фитоценология. – СПб.: С.-Петербургского ун-та, 1997. - 316с.
13. Колесникова Н.А. Школьный учебно-опытный участок-лаборатория исследовательской работы учащихся. – Тобольск: изд. ТГПИ, 2003. – 61 с.
14. Крылов А.Г. Жизненные формы лесных фитоценозов. – СПб.: Наука, 1984. – 189с.
15. Лакин Г.Ф. Биометрия: уч.пособие. – 4-е изд., перераб. И доп. – М.: Высшая школа.1990. – 351с.
16. Лотова Л.И. Морфология и анатомия высших растений: Учебное пособие. – Киев: Эдиториал, 2000.
17. Мазная Е.А. Влияние промышленных выбросов на состояние и структуру ценопопуляций Vaccinium myrtillus L. И Vaccinium vitis-idaea L. (Кольский п-ов) //. Растительные ресурсы. – 2001. – Т.37вып.3.
18. Медведев С.С. Физиология растений. – СПб., 2004. – 336 с.
19. Минаева В.Г. Лекарственные растения Сибири. – Новосибирск: Наука, 1991. – 496 с.
20. Пояркова А.И. Ericaceae // Флора СССР. Т.18 – М. – Л., 1952. – С. 22-107.
21. Работнов Т.А. Луговедение. – М.: МГУ, 1984. – 319с.
22. Работнов Т.А. Фитоценология. – М.: МГУ, 1978. – 383с.
23. Рябицев В.К., Плотников В.В., Смирнов Н.Г., Ольшванг В.Н., Богданов В.Д., Мухин В.А., Троценко Г.В., Пешкова Н.В. Природа Тюменского Севера. – Свердловск: Средне-уральское книжное издательство, 1991. – 432 с.
24. Смирнова О.В. Структура травяного покрова широколиственных лесов. – М.:Наука, 1987. – 205с.
25. Сокол А.П. Экологические шкалы болотных растений // Взаимосвязи компонентов лесных и болотных экосистем средней тайги Приуралья. – СПб.: Наука, 1980. = С.230-240.
26. Толмачев А.И. К истории возникновения и развития темнохвойной тайги. – М.: АН СССР, 1954.
Приложение 1
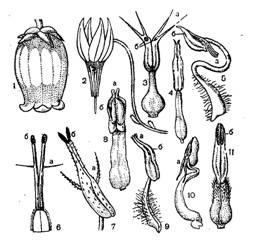
Рис.1. Цветки и тычинки вересковых
Цветки: 1 – хамедафна болотная (Chamaedaphne calyculata); 2 – клюква (Oxycoccus) Тычинки: 3 – зеновия припудренная (Zenobia pulverulenta); 4 – хамедафна болотная (Chamaedaphne calyculata); 5 – лиония кустарниковая (Lyonia fruticosa); 6 – черника тычиночная (Vaccinium stamineum); 7 – черника обыкновенная (Vaccinium myrtillis); 8 – эллиотия кистевидная (Elliotia racemosa); 9 – гаультерия лежачая (Gaultheria procumbens); 10 – пиерис пышноцветный (Pieris floribunda); 11 – оксидендрум древовидный (Oxydendrum arboretum): а- тычиночные придатки; б- поры.
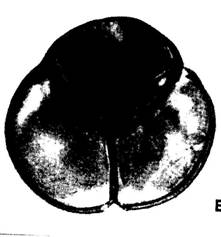
Рис.2. Пыльцевые зерна цветков растений семейства вересковых