
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Рефераты по косметологии
Рефераты по криминалистике
Рефераты по криминологии
Рефераты по науке и технике
Рефераты по кулинарии
Рефераты по культурологии
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Рефераты по косметологии
Рефераты по криминалистике
Рефераты по криминологии
Рефераты по науке и технике
Рефераты по кулинарии
Рефераты по культурологии
Курсовая работа: Біологічний контроль збудників хвороб рослин
Курсовая работа: Біологічний контроль збудників хвороб рослин
ЗмістВСТУП
РОЗДІЛ 1. Сучасний стан епідеміології вірусів вищих рослин
РОЗДІЛ 2. Шляхи розповсюдження вірусів рослин в природі та роль факторів навколишнього середовища
2.1 Клімат та погодні умови
2.2 Розповсюдженість вірусів. Біотичні фактори
2.3 Розповсюдженість вірусів. Абіотичні фактори
2.4 Шляхи передачі вірусів рослин в природних умовах
Розділ 3. Найбільш поширені вірусні захворювання серед вищих рослин
3.1 Кількісна характеристика та класифікація вірусів рослин
3.2 Віруси, які вражають певні види рослин
3.3 Віруси, які уражують широке коло рослин господарів
3.4 Боротьба із вірусними хворобами рослин
РОЗДІЛ 4. Біологічний метод: його суть та використання біопрепаратів
4.1 Біологічний метод
4.2 Ефективне застосування біопрепаратів при вирощуванні овочевих культур
4.3 Бактеріальні препарати і біогумат та їхні властивості
ПРАКТИЧНА ЧАСТИНА
Аналіз способів застосування бактеріальних препаратів і регулятора росту біогумату на овочевих культурах
Ефективність використання бактеріальних препаратів і регулятора росту біогумату при вирощуванні овочевих культур
Застосування бактеріальних препаратів і регулятора росту біогумату на помідорах
ВИСНОВКИ
СПИСОК ВИКОРИСТАНОЇ ЛІТЕРАТУРИ
Вступ
Актуальність. Протягом останнього часу в фітовірусологи, як і в інших молекулярно-біологічних дисциплінах, спостерігається значний інтерес до розвитку досліджень на популяційному та еколого-популяційному рівні, і ця тенденція є загальною для багатьох традиційних молекулярно-біологічних досліджень. Значні успіхи у фітовірусологи на молекулярно-біологічному рівні, такі як розшифровка структури генів, білок-білкової та білок-НК взаємодії, розшифровка АК-послідовностей та тривимірної будови структурних та регуляторних вірусних білків та багато іншого контрастує з нашими знаннями відносно поведінки вірусів рослин в природніх умовах біоцеозів. Поява в останні десятиріччя нового покоління діагностичних методів для визначення наявності фітовірусів (таких як ІФА, RIA, RIPA, ПЛР, ЛЛР, та ін) дали змогу на новому рівні вивчати розповсюдженість фітовірусів в навколишньому середовищі. Актуальність даного питання полягає в його як фундаментальності, так і в практичному значенні. Визначення ареалу розповсюдженості, механізмів передачі, кола рослин-господарів, реакції на зміни оточуючого середовища дає можливість не тільки більш повно характеризувати того чи іншого представника царства Vira, але і дасть змогу прогнозувати появу та розвиток вірусних хвороб і правильно виробляти стратегію і тактику боротьби з ними — запровадження стійких сортів, цілеспрямована боротьба з резерватами та переносниками, отримання безвірусного посадкового матеріалу.
Перспективним напрямком є моделювання розвитку хвороб, однак для вірусних хвороб рослин адекватні моделі практично відсутні, на відміну від фітопатогенних грибів та мікроорганізмів.
Встановлення можливих взаємозв'язків між наявністю вірусних антигенів в рослинах та ґрунті є дуже важливим у розумінні розповсюдження фітовірусів рослин та циркуляції їх в агроценозах.
Предмет дослідження – збудники хвороб рослин.
Об`єкт дослідження – застосування біологічного контролю збудників хвороб рослин.
Мета роботи – з`ясувати шляхи проникнення вірусних інфекцій в рослинні організми
Основні завдання:
1) охарактеризувати сучасний стан епідеміології вірусних захворювань рослин на Україні;
2) розглянути шляхи розповсюдження вірусів рослин в природі та роль екофакторів у цьому процесі;
3) дати стислу характеристику найбільш поширених в Україні вірусів рослин;
4) розглянути основні заходи боротьби з вірусами рослин;
5) проаналізувати суть та перспективи застосування біологічного контролю збудників рослин.
Розділ 1. Сучасний стан епідеміології вірусів вищих рослин
На порозі двадцять першого століття епідемії хвороб рослин все-таки призводять до значних втрат у продовольстві, промисловості, лісових та сільськогосподарських культурах всюди у світі. Населення нашої планети продовжує збільшуватись і якщо ми хочемо мати достатньо продовольства, одягу та помешкань для поточного та майбутніх поколінь, важливим є зменшення або й зовсім усунення втрат від хвороб рослин.
Епідеміологія хвороб рослин являє собою вивчення просторових та часових змін, що відбуваються під час епідемій хвороб рослин, які викликані популяцією патогенів у популяції рослин. Епідемія трапляється, коли відбуваються зміни у інтенсивності захворюваності серед популяції господаря у часі та просторі. Патогеном можуть бути гриби, бактерії, мікоплазмоподібні організми (МПО), віруси, віроїди, нематоди або паразитичні рослини. Рослини можуть бути частиною будь-якої сільськогосподарської, садової, лісової, або природної екосистеми. Взаємодія патогена та рослини-господаря, що призводить до розвитку хвороби, може трапитися у будь-якому наземному, водному, або штучно створеному середовищі. Сума усіх патогенів, господарів та середовищних компонентів являє собою рослинну патосистему.
Наука, що вивчає епідемії хвороб рослин (тобто, епідеміологія) має як описову, так і кількісну фази. Спочатку ставиться питання, наприклад, де епідемії трапляються, коли можуть епідемії очікуватися, які фактори відіграють роль у розвитку хвороби. Коли знання патосистеми збільшується, починаються питання більш кількісної природи, наприклад, скільки інфекційних одиниць патогену потребується, щоб започаткувати епідемію, скільки хвороба триває, як швидко буде хвороба розвиватися, та як далеко може інфекційна одиниця патогену розповсюдитись. Щоб відповісти на ці питання, використовується експериментальна та аналітична техніка класичної фітопатології та техніки, розроблені у агрономії, ботаніці, хімії, екології, ентомології, генетиці, математиці, метеорології, фізиці та статистиці.
Віруси та мікоплазмоподібні організми (МПО) рідко є окремими, незалежними інфекційними одиницями в оточуючому середовищі. Навіть частки цих патогенів звичайно пов'язані з рослиною-господарем або з переносниками (комахами чи нематодами). Таким чином, визначення рівня інфекційних одиниць в першу чергу робиться шляхом оцінки титру патогена в рослинах-господарях, або шляхом моніторингу популяції відомих переносників та визначення частки популяції переносника, що несе патоген рослин. У випадку титру вірусу точну кількість його не вимірюють, тоді як комаха або нема-індії, кожна з яких є переносником патогену, насправді є інфекційною одиницею або пропагулою епідемічного розвитку.
Зібрані зразки з інфекційними збудниками вірусної природи можна ідентифікувати за допомогою електронного мікроскопу, візуального мікроскопічного спостереження або шляхом біологічного тестування. Біохімічні технології можуть бути корисними для вірусів та МПО. При виборі методу моніторингу потрібно приймати до уваги складність ідентифікації, необхідний час та вартість витратних матеріалів.
Концепція збудника не зовсім чітка для вірусів та МПО. Хоча існують окремі частки або одноклітинні організми, вони мало впливають на епідеміологію хвороб, спричинених вірусами або МПО. Для багатьох з цих хвороб патоген передається з рослини на рослину за допомогою діяльності комах. Деякі ґрунтові віруси переносять нематоди або гриби. Обчислення кількості інфекційних одиниць поза рослиною-господарем роблять за допомогою збирання зразків векторів. Обчислення вірусів та МПО в їхніх рослинах-господарях для епідеміологічних завдань дає кращі результати при оцінці інтенсивності хвороби.
Хоча рослинні патогени переносять різні ряди комах, найбільш численні вектори має ряд Homoptera. Родини цього ряду включають горбаток, білокрилок, кокцид та попелиць. Взагалі, комахи Homoptera переносять 241 вірус та 45 МПО. Як і у випадку з повітряними грибами, комах можливо обчислити за їхнім накопиченням (зіткненням) із стаціонарними пастками, або їх концентрації в повітрі за допомогою всмоктувальних пасток. Далі, можливо визначити кількість особин, що несуть патоген та здатні до його трансмісії.
Оцінка хвороб рослин є одним з найбільш важливих та часто найбільш складних завдань у епідеміології хвороб рослин. Оцінка хвороби на даний час є підвалиною статистичного аналізу даних, зусиль з моделювання та інтерпретації патосистеми. Похибка, введена під час оцінки хвороби, може бути дуже малою на початку, але збільшитись при подальших епідеміологічних аналізах. Отже, ступінь впевненості у висновках та спроможність розрізнювати справжні ефекти не буде більшим, ніж рівень надійності при оцінці хвороби.
Оцінка хвороби звичайно триває багато часу і тому відносно дорога. Кропітке планування — як, коли, де та ким — хворобу буде оцінено, дуже істотне. Планування вимагає ясної концепції мети специфічного дослідження та докладне знання патосистеми.
Оцінки хвороби можуть бути кількісними, якісними, або комбінацією цих двох типів. Кількість наявної хвороби можна назвати поширеністю хвороби. До інтенсивності хвороби входять розповсюдженість хвороби та суворість хвороби. Додатково, такі характеристики, як концентрація або титр пропагативних або інфекційних одиниць (віріонів) у тканинах, можуть віддзеркалити інтенсивність хвороби. Якісні оцінки хвороби можуть включати як ринкову якість (вміст протеїну, олії, або вміст поживних речовин у господарі або Його продукті) так і такі фактори, як поживність, смак та запах.
Поширеність хвороби — це число одиниць рослин, що візуально уражено, звичайно до загального числа одиниць, що оцінено. Якщо розглядати рослину в цілому як одиницю, тоді поширеність хвороби є пропорція або процент рослин із симптомами хвороби. Поширеність хвороби може також бути оцінена, використовуючи окремі гілки, листки або коріння як рослинну одиницю.
Поширеність та суворість як виміри оцінки вживають разом, і правомірність кожного виміру повинна бути встановлена для кожної окремої патосистеми. Поширеність часто найбільш легка та найбільш швидка міра хвороби. Число зів'ялих рослин томату, почорнілі колосся пшениці, корінь із пухлинами або листя з некрозами чи мозаїчними симптомами можуть бути пораховані досить точно та акуратно. Звичайно поширеність є відповідною мірою хвороби для патосистем, у котрих один некроз на одиницю рослини є фатальним, також як і у випадку з хворобами в'янення та багатьма системними вірусними хворобами. Поширеність симптоматичних листків aбo гілок може також бути мірою хвороби для комерційних плодових дерев, тіньових лісів, тощо. Оцінка або вимірювання суворості хвороби часто більш тяжке та відбирає більше часу. Також, точність та відтворюваність може бути меншою, ніж у випадку поширеності. Суворість, однак, може бути більш важливою та корисною мірою дослідження хвороби для багатьох патосистем, що мають відношення до управління хворобами, динаміки чисельності та оцінки втрат врожаїв.
У більшості випадків, суворість хвороби розглядають як частку розглянутих рослинних одиниціь. Свідчення з фізіології рослин та досліджень втрат врожаїв показують, що врожай рослин може бути визначений із загальної суми уражених областей або, більш точно, із загальної суми здорових областей. Це примушує епідеміологів винайти шляхи для оцінки ступеню абсолютної суворості хвороби, хоча необхідно оцінити втрати врожаю. Однак на сьогоднішній день можуть бути застосовані декілька різноманітних шляхів репрезентації інтенсивності.
Терміни та частоти оцінок хвороби визначаються пагосистемою та метою оцінки. Оцінки можуть ґрунтуватися на календарних строках, фізіологічно-середовищних чинниках, або в залежності від стадій росту. Календарна шкала є найбільш часто використаною шкалою часу у епідеміологічних дослідженнях, що часто проводять через 5-, 7-, 10-, або 30-денні інтервали. Календарні оцінки є зручними та дозволяють скласти регулярний розклад роботи. Такі оцінки також забезпечують регулярно розмежовані дані для аналізу прогресу хвороби, у якому час використаний як незалежна змінна.
Фізіологічно-середовищна шкала часу використовує середовищні властивості або комбінації властивостей, що є важливими для фізіологічного стану господаря або патогену. Цей часовий масштаб ґрунтується на концепції, що певне число середовищних одиниць повинне бути акумульовано для того, щоб ключова фізіологічна подія трапилася. Число днів з певною температурою, вищою за мінімальну — приклад цього типу шкали часу. Цей тип шкали вимагає безперервного моніторингу змінних середовища (або доступ до таких даних). Складання розкладу у такому випадку оцінки дещо більш важке, ніж з календарною шкалою, якщо бажані регулярні інтервали одиниць. Такими шкалами користувались відносно рідко в фітопатології; однак, приклади є, та й подальше дослідження переваг цієї шкали часу виправдане.
Розширення фізіологічно-середовищної шкали базується на стадіях росту господаря. В межах цієї пов'язаної із господарем шкали, рослина сама слугує як інтегратор всіх середовищних ефектів. Припускаючи, що стадії росту ґрунтуються на ключових подіях у життєвому циклі господаря, оцінки хвороби при цих ключових стадіях забезпечують інформацію, що може бути корисною для аналітичної або модельної мети, для оцінки втрат врожаю, або для оцінки матеріалу у програмах вирощування рослин, стійких до хвороб.
Частота оцінок може залежати від частоти подій, вимог щодо інформації про різні стадії епідемії, або економічних міркувань. На фізіологічно-середовищній або шкалі стадій росту, частота специфічних подій (наприклад, акумуляція температурних днів або перехід до наступної стадії росту) визначить частоту оцінок хвороби. Якщо потрібен докладний статистичний або математичний аналіз епідемії, частота оцінки може визначатись потребою в специфічній кількості спостережень або потребі в спостереженнях під час "критичних" частин епідемії. Для епідемії, що триває 90 днів, тижневі оцінки дадуть 12-13 рівномірно розподілених спостережень. Ця кількість певно є достатньою для більшості випадків; однак, якщо економічні міркування диктують, що можна зробити менше ніж 12 спостережень, буде більш сприятливим зробити оцінки з нерегулярними інтервалами.
Економічні міркування при оцінці хвороби повинні бути врівноважені з потребою у специфічній інформації. Якщо час є обмежувальним фактором (як це і є звичайно), тоді необхідно визначити мінімальне число оцінок. Мета оцінки та знання джерел змін в межах та серед експериментальних одиниць буде диктувати, чи краще зробити більше спостережень у одному місці, або менше спостережень у багатьох місцях.
Можна зробити висновок про те, що маючи досить ґрунтовну теоретичну базу, ми ще не маємо більш-менш стандартних методик щодо об'єктивної оцінки розвитку вірусних інфекцій в агроценозах. Цей факт свідчить про актуальность цієї проблеми як її вузькому (вірусологічному) так і в широкому (сільськогосподарському та екологічному) розуміннях.
Розділ 2. Шляхи розповсюдження вірусів рослин в природі та роль факторів навколишнього середовища
2.1 Клімат та погодні умови
Віруси рослин мають здатність досить швидко розповсюджуватися в біоценозах. Вони можуть поширюватися завдяки переносникам (кліщі, попелиці, нематоди, гриби), передаватися при вегетативному розмноженні через бульби, живці, цибулини. Більшість фітовірусів передаються щепленим. Близько 90 різних вірусів рослин передаються насінням, деякі фітовіруси можуть викликати інфекцію без участі переносників, коли збудник знаходиться в грунті. Знаній екології збудника необхідне для використання необхідних засобів боротьбі проти нього (рис. 1.).
При ураженні рослин — господарів фітовірусами найчастіше розвивається системна реакція. При цьому інфекція часто протікає з яскраво вираженими симптомами у вигляді мозаїки, скручування, хлорозу, появи енацій та ін. Така інфекція може закінчуватись навіть загибеллю інфікованих частин рослин або і всієї рослини. Разом з тим, інколи перебіг інфекційного процесу проходить без чітких ознак хвороби, які помітні за габітусом рослин. Часто уражений організм може бути прихованим носієм інфекції. Продуктивність такі(Х рослин знижується в незначній мірі. Так реагують на вірусну інфекцію рослини-резерватори, переважна більшість яких відноситься до бур'янів.
Бур'яни (щириця біла, гірчиця біла, суріпиця звичайна, пирій повзучий та ін.) займають особливе місце серед факторів, що знижують врожай сільськогосподарських культур. Для цих рослий характерні велика енергія насіннєвого та вегетативної о розмноження, тривале зберігання схожості насіння. Вони сильно виснажують та висушують грунти, часто являються джерелом розповсюдження хвороб та шкідників сільськогосподарських культур. Джерелом вірусів служить також насіння бур'янів. Завдяки його достиганню та обсипанню до збору сільськогосподарських культур, насіння накопичується на полях у великій кількості. Так, в одному шарі товщиною 30 см знаходиться в середньому від 100 млн. до 4 млрд. і більше насіння бур'янів на 1 га. При обробці грунтів значна їх частина переміщується в верхній горизонт, і їх кількість тут в сотні і тисячі разів перевищує кількість насіння, що висівається. Крім того, грунт може бути забрудненшій кореневищами бур'янів, що також можуть бути уражені вірусами.
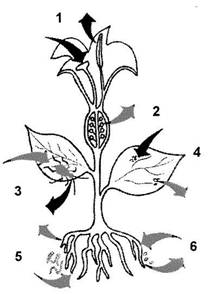
Рис. 1. Основні шляхи передачі фітовірусів.
1 - передача за допомогою пилку; 2 - передача за допомогою насіння; 3 - за допомогою векторів-комах; 4 - при прямому контакті з рослинами (механічному пошкодженні); 5 - за допомогою еекпюрів-нематод; 6 - за допомогою векторів-грибів.
Так, вірус мозаїки цукрового буряку уражує також щирицю, осот, кормові боби. Віруси, що викликають жовтяницю буряка, можуть перезимовувати в багатьох розповсюджених бур'янах.
Багато вірусів швидко мігрують завдяки переносникам. Переносниками можуть бути попелиці, цикадки, кліщі, нематоди, гриби та ін. Напевно, перше місце по передачі вірусів серед комах займають попелиці. Такі віруси, як вірус огіркової мозаїки, вірус жовтої карликовості ячменю, У-вирус картоплі, вірус мозаїки цукрового буряку, вірус жовтяниці цукрового буряку можуть передаватися різними видами попелиць. В деяких випадках, наприклад, між вірусом жовтої карликовості ячменю та попелицями, існує симбіотичний взаємозв'язок. При живленні попелиць на уражених цим вірусом рослинах підвищується їх плодовитість. Активними переносниками вірусу смугастої мозаїки пшениці є кліщі Aceria tulipae Keit та Асегіа tritici Schev. ВСМП може довгий час циркулювати в природі. Зимує вірус в інфікованих рослинах і кліщах. З весни та до збору врожаю кліщі інтенсивно розмножуються на рослинах, а потім мігрують на бур'яни (пирій повзучий, стоколос безостий та ін) і кукурудзу. Восени воші мігрують на сходи озимих та уражують їх вірусом (40).
Попелиці і кліщі, як і інші летючі комахи, є одними з важливих факторів, що визначають дальність розповсюдження віруса від джерела. Так, за один день попелиці відносяться вітром на десятки і навіть сотні кілометрів.
Грунт є також одним із джерел інфікування рослин вірусами. Потрапляючи в грунт, віруси здатні інфікувати нові рослини. Існує три групи вірусів, що передаються через грунт:
1. віруси, що передаються грибами;
2. віруси, що передаються нематодами;
3. віруси, переносник яких невідомий.
Крім вірусів, що переносяться або нематодами з родів Longidorus, Xiphinema і Trichodorus, або зооспорами ґрунтових грибів з родів Polymixa і Olpidium, існує група ґрунтових вірусів, представники якої "вільними" вібріонами зустрічаються в грунті і воді та можуть переходити без допомоги переносників від хворих до здорових рослин (non-vectored viruses), хоча деякі з них можуть переноситися нематодами або грибами.
Незважаючи на свою різноманітність, грунтові віруси мають ряд спільних ознак (за винятком ВОМ):
- не розповсюджуються літаючими переносниками;
- накопчуються в рослинах-господарях у високих концентраціях і виділяються через корені інфікованих рослин в грунт;
- надзвичайно стійкі поза живою клітиною;
- можуть інфікувати здорові рослини через корені без допомоги переносників;
- мають, як правило, широке коло рослин-господарів;
- більшість з них мають простий однокомпонентний геном;
- деякі з них можуть розповсюджуватись за допомогою безпосереднього контакту.
В грунт віруси рослин можуть потрапити як прямим (з листових, стеблових і кореневих залишків уражених рослин, та при віддачі вірусів коренями інфікованих рослин), так і непрямим шляхом (вірусовмісні екскременти та вимивання вірусів з контамінованих ґрунтів дощовими водами). Так, вірус огіркової мозаїки може проходити через травний канал людини або тварин, а потім зі стічними водами потрапляти в грунт і вражати рослини.
Джерелами контамінації "вільних" від вірусів ґрунтів або інших субстратів може бути, ймовірно, невелика кількість рослин, що постійно виділяють через корені вірус в навколишнє середовище. Деякі віруси настільки призвичилися до цього способу розповсюдження, що інфікують тільки кореневу систему рослин-господарів або окремі частини їх надземних органів. Діагностика в такому випадку ускладнена, і можна припустити, що віруси цієї групи розповсюджені більш широко, ніж думали до останнього часу.
На даний період в літературі описані 11 вірусів, у яких встановлена віддача вірусів через корені і 11 вірусів, у яких показана інфекція рослин через корені без переносників.
Численні віруси рослин містяться в ґрунті і в поверхневій воді, особливо в сільськогосподарських областях. Ступінь адсорбції і тривалість періоду, на протязі якого інфекційність зберігається і який може тривати тижні, а іноді і місяці, залежить від захисту вірусу і факторів зовнішнього середовища. Серед останніх це тип ґрунту, рівень вологості, рН, температура, вміст органічних субстанцій і звичайно зв'язана з цим біологічна активність середовища. Багато вірусів стабільні і здатні інфікувати рослини без допомоги векторів, а саме нематод та розповсюджених в ґрунті зооспор грибів. Передача вірусу з ґрунту та води ймовірно можлива через непошкоджені корені рослин. Віруси, які є в струмках, можуть переноситись на великі відстані. Іноді нові віруси, не знайдені у рослинах-господарях, були ізольовані з води та ґрунту.
До теперішньої о часу приблизно 9 вірусів були ідентифіковані і кілька інших ізольовані з ґрунту і води лісових екосистем. Вони головним чином належать до: Potex,- Tombus-, Tobamo-, Poty- та Necrovirus груп. Крім цього, ці віруси та принаймні 7 представників інших таксономічних груп, особливо Nepo- та Ilarviruses, були знайдені в деревовидних лісових рослинах. Такі збудники вірусних інфекцій можуть бути більш або менш шкідливі для інших видів рослин. Також багато вірусів було ідентифіковано в квіткових лісових рослинах. Ці патогени являють собою потенційну загрозу для економіки лісового господарства.
Невелика кількість досліджень поширення вірусів в цих субстратах (ґрунті та воді) спонукає нас проводити роботу по ідентифікації та розповсюдженості вірусних інфекцій в агроценозах України.
Останнім часом увага дослідників була привернута до вірусних систем збереження, як видно з екологічних робіт. Для деяких вірусів добре розвинена здатність до збереження на полі (дільниці) компенсується обмеженою здатністю до поширення на нові дільниці. Навпаки, інші віруси, особливо деякі з тих, що передаються персистентим способом повітряними векторами, широко передаються на нові дільниці, але не здатні до довгого збереження на даній дільниці (якщо мова йде виключно про агроценоз). Володіння ефективними способами як передачі, так і збереження, є характерною рисою деяких з більшості постійно циркулюючих вірусів.
Тому що віруси з однієї таксономічної групи мають подібну систему виживання, а різні типи рослинних угрупувань ймовірно мають схильність до різних видів систем виживання, можна очікувати, що специфічні групи вірусів будуть краще адаптовані до специфічних рослинних угрупувань. В середовищі, придатному для контактної передачі тобамо- та потексвірусів, рослини виробляють високу концентрацію відносно стабільних часточок в їхніх господарях. Інші адаптовані до культивованих рослин віруси (CULPAD viruses), включають іларвіруси, які головним чином можна знайти в дерев'янистих видах. Вони передаються пилком при запиленні рослин. Віруси, які переносяться за допомогою грибів (групи вірусу жовтої мозаїки ячменю), персистують на рестингспорах їхніх векторів і віддають перевагу монокультурам. З іншого боку, є віруси, адаптовані до диких рослин (WILPAD viruses) які включають багато груп, таких як непо-, тобра-, геміні-, лютеовіруси. Ці віруси мають широкий спектр господарів і довготривалу персистенцію на їхніх векторах. Ці властивості підходять до виживання в угрупуваннях, які складають багато видів рослин.
Екологія вірусів рослин, а віднині і епідеміологія вірусних хвороб рослин не повинна розглядатись як відбір незмінних зразків, вони можуть змінюватись під впливом багатьох факторів, таких як довгострокова зміна погоди, зміни в методах вирощування рослин, зміни екологічних параметрів ценозу. Для прикладу, вирощування рослин з використанням напівпрозорих пластикових листів в деяких країнах привело до поширення вірусів на захищених полях, а в Англії зростання заміщення ярого ячменя на озимий супроводжується зростанням кількості і поширенням вірусів, які є проблемними для цих культур.
Крім того, до цих "сторонніх" змін в методах культивування сільськогосподарських рослин є більш прямі наслідки від спроб контролювати віруси. Наприклад, вирощування культури томатів, що мають ген стійкості до ВТМ (Тm-1), призводять до селекції і поширення нестійких штамів, але ці штами, очевидно, мають лімітовану біологічну придатність, і коли тиск, нав'язаний стійкою культурою, знімається, вони зменшуються в поширенні.
Найімовірніше, віруси рослин протидіють стійкості, що контролюється одним чи двома генами господаря, більш повільно, або не контролюються взагалі. Це очевидно відбувається через те, що їхні геноми мають обмежену ємність кодів і їхній генний продукт є багатофункціональним, або через те, що їхні засоби передачі обмежують ступінь, до якого рослини з стійким генотипом піддаються інокулюму. В цій "боротьбі генів" нестійкі варіанти можуть з'являтися, але як вважається, можуть бути відносно невдалими. Протилежними в цій взаємодії є селективні ефекти дії вірусів на їхніх господарів, які звичайно проявляються після багатьох років. Таким чином, гени для толерантності або резистентності до інфекції звичайно знаходять на культивованих посівах на ділянках, де рослини постійно або регулярно піддаються вірусній інфекції довгий проміжок часу — резистентність кукурудзи до вірусу смугастості кукурудзи в Ісландії, резистентність картоплі до декількох вірусів в Південній Америці є тому прикладом.
В епідеміології географічний діапазон факторів, які працюють, є дуже важливим, тому шкала подій збільшується, що приводить до більшої складності системи. Є багато прикладів того, що можна домогтися при детальному вивченні розповсюдженості вірусів на специфічних ділянках. Важкі для розуміння з точки зору епідеміології вірусу великі континентальні земляні маси, де фактори, діючі на протязі сотень кілометрів, можуть привести до значних ефектів, і де щорічна сфера дії інфекції може піддаватися сильним флуктуаціям. В таких областях останні досягнення методів, заснованих на вивченні великомасштабних рухів повітряних мас, можуть частково принести успіх для картографії міграції повітряних векторів. Дійсно, такий спосіб вивчення впливу однієї частини континенту на іншій показаний на прикладі впливу великомасштабного переміщення вірусу коричневої плямистості рису.
Для аналізу причин і перебігів епідемій останнім часом з'явилося багато корисних методик. Велика кількість рослин може зараз бути протестована на присутність вірусної інфекції з допомогою чутливих серологічних методів, таких як ELISA. Досягнуто прогресу в детекції вірусів серологічними методами в індивідуальних векторних організмах, зібраних на полях, та різні генотипи вірусів можуть бути вивчені з допомогою гібрідізації комплементарної ДНК. Такою є робота по порівняльній екології різних штамів вірусів. Можна очікувати, що комп'ютерний аналіз може допомогти розібратися з великими масивами даних, одержаних після різнобічного вивчення і постійних записів стану зовнішнього середовища, а також допомогти зробити життєво необхідний висновок по поясненню подій та їх прогнозуванню.
Ситуація, що існує в дійсності по розвитку кожного вірусу в конкретній місцевості або в світовому масштабі, є результатом складної взаємодії багатьох фізичних та біологічних факторів. Розуміння екології вірусу щодо окремої культури та конкретної місцевості важливе для розробки відповідних методів боротьби із захворюваннями, що він викликає. Як і по відношенню до більшості інших облігатних паразитів, основними екологічними факторами, що слід розглядати, звичайно вважають шляхи передачі вірусів від рослини до рослини, а також шляхи, якими інші фактори впливають на розповсюдженість вірусу.
2.2 Розповсюдженість вірусів. Біотичні фактори
Вірус, що зберігає стабільність структури, знаходячись як всередині рослини, так і зовні і досягає високих концентрацій в тканинах, має, зрозуміло, більшу ймовірність вижити та розповсюдитись, ніж той, що цими властивостями не володіє. Наприклад, ВТМ може на протязі довгого періоду зберігатись в мертвому рослинному матеріалі, що знаходиться в грунті, котрий в такому разі стає джерелом інфекції для наступних культур.
Віруси або штами вірусів, що переміщуються по рослині повільно від того місця, де пройшло ураження, мають меншу ймовірність вижити і потім ефективно розповсюджуватись, ніж ті з них, що транспортуються швидко. Швидкість руху вірусів відіграє певну роль, якщо її розглядати з точки зору тривалості життя окремих рослин-господарів. Так, віруси, що уражують довгоживучі дерева та кущі, можуть рухатись по своїм рослинам-господарям значно повільніше, ніж ті, що інфікують однорічні рослини. Віруси, що можуть проникати в насіння і зберігатись там, мають суттєві переваги порівняно з іншими, з точки зору розповсюдження, а також здатності до виживання. Розповсюдження вірусу некрозу тютюну у більшості рослин-господарів обмежене кореневою системою, і існування цього вірусу в природі залежить від здатності грибів-переносників передавати його через грунт і уражувати таким чином інші рослини. Ймовірність виживання вірусу, що викликає швидко-прогресуюче системне захворювання, яке призводить до загибелі рослини, значно менша, ніж у тому випадку, коли вірус викликає у рослин слабкі або середньої тяжкості захворювання, при яких вони продовжують вегетувати та розмножуватись. Цикадки інфікуються спочатку штамами вірусу кучерявості верхівки цукрового буряку, що викликають слабкі симптоми на рослині-господарі. Вірулентні штами цього вірусу викликають загибель рослин до того, як на них закінчить, розвиток наступне покоління цикадок. Таким чином, в деяких умовах вірулентні штами мають тенденцію до самознищення. В країнах з високими температурами влітку може проходити інактивація вірусів in vivo в природних умовах. Так, в деяких районах Індії бульби картоплі, що зберігаються на звичайних складах, можуть звільнитись від вірусу згортання листків, в той час як бульби, що зберігаються в охолоджених сховищах, залишаються повністю інфекційними. Аналогічним чином рослини суниці звільнялись від вірусу зморшкуватості.
Таким чином основні фактори, що впливають на виживання штаму, такі:
- ефективна передача комахами або іншими засобами;
- більш швидке розмноження і більший врожай, ніж у конкуруючих штамів;
- розвиток слабкої або не надто сильної форми захворювання. Віруси широко варіюють по відношенню до спектру тих видів рослин-господарів, що вони уражують. Так, деякі віруси, що уражують полуниці, обмежені родом Fragaria. А вірус бронзовості томатів має дуже широке коло господарів, до якого входять як однодольні, так і дводольні. Відомо, що цей вірус уражає рослини 166 видів, що входять до 34 родин.
Віруси з дуже вузьким колом рослин-господарів, ймовірно, виживають тому, що ці рослини багаторічні, або розмножуються вегетативно, або внаслідок того, що такі віруси передаються насінням. Для багатьох нематод та вірусів, які вони переносять, характерний широкий спектр рослин-господарів, до яких входять і багаторічні деревоподібні рослини.
Оцінюючи віруси рослин в цілому, вважається, що найважливішим агентом розповсюдження, а таким чином, і виживання вірусів є літаючі комахи-переносники, що живляться соком рослин, зокрема попелиці. Шлях розповсюдження вірусу в межах окремої культури, а також швидкість та відстань, на яку його може бути перенесено, залежить від багатьох факторів (віруси, що передаються через повітряне середовище):
- джерело інокулюму; інокулюй може потрапити в культуру зовні, від хворих рослин в самій культурі, що мали вірус при передачі насінням або при вегетативному розмноженні, а також від
бур'янів та інших рослин, що знаходяться в цій культурі;
- кількість потенційно інфекційного інокулюму;
- природа та modus vivendi переносника (наприклад, щодо попелиць, то має значення чи це колонії, чи крилаті форми);
- передається вірус за допомогою стилету, є він циркулюючим чи розмножується в переноснику;
- час, коли переносник стає активним, відносно строків вегетації рослин даної культури;
- погодні умови.
Ван Дер Планк ще у 1946 році запропонував метод, за допомогою якого можна визначити, чи розповсюджується вірус в межах ділянки від ураженої рослини, чи він потрапляє зовні. Його засновано на припущенні, що вірус, який потрапляє зовні, буде викликати випадкове ураження рослин, тоді як при його розповсюдженні від хворої рослини до хворої, уражені рослини будуть розташовані групами. Таким чином, оцінюючи кількість пар уражених рослин відносно очікуваної, роблять висновок про походження вірусу.
Для вірусів, що розповсюджуються через грунт швидкість розповсюдження залежить від:
- типу переносника;
- структури, вологості, щільності і т.п. ґрунту;
- густини рослин-господарів;
- кліматичних умов;
- інших факторів.
Однак, відомі і виключення. Так, ВТМ — один з небагатьох вірусів, що в помітних кількостях поширюється через грунт без допомоги переносників.
Окрему групу феноменів складає перенесення вірусів на великі відстані. Більшість з таких явищ спонтанні і здебільшого не підлягають передбаченню. Вони порушують усталену рівновагу в ценозі, і тому довгострокові прогнози можливі лише на відносно стабільних системах, оскільки охоплюють коло тільки відомих характеристик.
На ураженість деякої культури захворюванням вірусної етіології значний вплив мають методи її вирощування та заходи обробки, що застосовують протягом багатьох сезонів. Тому будь-який прогноз має містити рекомендації щодо вдалого сівообігу, засобів обробки, термінів висіву та збору рослин. Вплив різного роду факторів, що мають значення у данному випадку, можна розглянути на таких прикладах.
Якщо поряд із полем озимини знаходиться поле ярої пшениці, ураженої вірусом смугастої мозаїки, то висів озимини до збору ярової пшениці призводить до великого ступеня її ураження ВСМП, який переноситься кліщами.
Тип сівообігу має деякий вплив на розповсюдженість вірусів, що здатні зимувати в бур'янах або у падалиці попередньої культури. В деяких культурах велика кількість спонтанно ростучих рослин, що містять вірус, може вегетувати довгий термін. Донкастер та Грегорі довели, що для того, щоб позбавитись від таких рослин картоплі, необхідно 5-6 років. А у випадку багаторічних культур кількість хворих рослин збільшується із віком культури.
На розповсюдженість та зберігання вірусів в грунті або рослинних залишках можуть мати вплив методи обробки грунту. Нематоди, а також гриби, що є переносниками вірусів, можуть розповсюджуватись при переміщенні грунту під час обробки. На зберігання ВТМ в рослинних залишках впливає ступінь аерації грунту, його вологість. Якщо попередньою культурою була картопля, то обробка грунту у холодну погоду може суттєво знизити виживання невикопаних бульб.
Щільність насадження та розмір рослин впливають на ймовірність їх зараження. Так випадки ураження вірусом жовтяниці буряку, вірусом мозаїки буряку та вірусом мозаїки кольорової капусти відмічались рідше, коли відстань між рядками рослин була менша. Також низькі рослини мають менше шансів бути відвіданими переносником, ніж високі.
Останнім часом все більшу увагу привертають бур'яни та дикоростучі рослини, роль яких в минулому недооцінювалась.
2.3 Розповсюдженість вірусів. Абіотичні фактори
При аналізі даних по захворюваності вірусними хворобами цукрового буряку в Англії було зроблено висновки, що сезонні коливання частоти ураження є наслідком довготривалого впливу погодних умов на популяцію попелиці-переносника протягом більшої частини сезону вегетації. Вони впливають на строки та чисельність попелиць, що мігрують на культуру, на розвиток популяції всередині культури та на її переміщення. Ґрунтові умови можуть різним чином впливати на ступінь ураження вірусними захворюваннями. На високо родючих ґрунтах ймовірність розповсюдження вірусних хвороб збільшується. Так, внесення органічних та деяких неорганічних добрив збільшувало на картопляних полях кількість рослин, що уражені вірусом скручування листків картоплі та УВК. На ступінь ураження, безперечно, впливає живлення рослин, пригнічуючи симптоми або роблячи їх більш чіткими. Як вже обговорювалось, ґрунтові умови відіграють помітну роль, впливаючи на зберігання ВТМ в рослинних залишках. Інактивація вірусу проходить швидше у вологих, добре аерованих ґрунтах, ніж у сухих, щільних або заболочених.
Таким чином, підсумовуючи, можна сказати, що не всім вірусам притаманні всі ті властивості, що сприяють їх виживанню та розповсюдженню, але кожному з них властива така їх комбінація, що дозволяє зберігатись в даних умовах більш-менш ефективно. Вважають, що при довготривалому впливі природного добору на вірус та генотип рослини-господаря і при відсутності втручання людини віруси зможуть так адаптуватися до рослин-господарів та своїх переносчиків, що їх дія буде мінімальною. Але, оскільки найближчим часом такого не передбачається, єдиним шляхом для людства є боротьба з вірусними захворюваннями, і не останнє місце в упередженні епіфітотій посідає їх прогнозування.
2.4 Шляхи передачі вірусів рослин в природних умовах
Термін життя окремої рослини обмежений, тому необхідною умовою виживання вірусу є перехід його від одної рослини до іншої. Джерелом інфекції може бути насіння або пилок, вегетативні частини рослини, бур'яни, комахи (у випадку персистентних вірусів), грунт. Передача вірусів за допомогою насіння представляє собою дуже ефективний спосіб ураження будь-якої культури на ранніх стадіях розвитку, так як на ділянці, де вирощується дана культура, з'являються хаотично розміщені осередки інфекції. Таким чином, якщо на даній ділянці вірус може передаватися від рослини до рослини іншими способами, внесення віруса в культуру за допомогою насіння являє собою важливий економічний фактор. Віруси таких родів, як Hordeivirus та Capillovirus, а також криптовіруси родини Partitiviridae передаються тільки насінням, але за допомогою насіння можуть передаватися і деякі представники Trichovirus, Carlavirus, Badnavirus, Machloraovirus, Sobemovirus, Idaeovirus, Tobamovirus, Closterovirus, Tombusviridae. Трансплантаційна передача вірусів може проходити за допомогою вегетативних частин рослини (пагонів, бульб, цибулин, вусиків), щепленням (привій, підвій) та за допомогою повитиці (Cuscuta spp.). Повитиця, що паразитує на вищих рослинах, належить до родини Convolvulaceae. Вона утворює гаусторії, які проникають у провідну систему росли-ни-господаря, і таким чином, може передавати віруси рослин (85). Трансплантаційна передача характерна для представників багатьох груп вірусів рослин — Trichovirus, Capillovirus, Closteroviridae, Badnavirus, Reoviridae, Tombusviridae. Механічний або контактний спосіб передачі вірусу має особливо важливе практичне значення на ранніх етапах росту польових культур. В природних умовах віруси можуть передаватися при контакті між собою, при застосуванні агротехнічних засобів. В останні роки велика кількість фітопатогенних вірусів, що не переносяться векторами, була виявлена в ґрунтах, ґрунтових водах, ріках, озерах та в морі. Рослинні патогени можуть різними методами попадати в грунт. Найбільш ймовірним є проникнення вірусів з інфікованими рослинами або рослинними залишками, в яких віруси можуть прямим або опосередкованим шляхом попадати в грунт. Прямий шлях полягає в тому, що інфіковані рослинні залишки залишаються в грунті (корінці) або з залишками рослин попадають в грунт (паростки, вегетативні органи). При цьому слід звернути увагу на те, що віруси, які містяться в рослинних залишках частіше довше залишаються іфекційними, ніж віруси, які знаходяться в грунті у вигляді вільних віріонів. Інфіковані вірусом рослини, які вживають люди та тварини, що можуть проходити через шлунок без втрати інфекційності та контамінвати грунт через екскременти, представляють непрямий шлях контамінації грунту. Представники Tombusviridae та Diantovirus передаються без допомоги векторів через грунт, хоча деякі віруси, наприклад, вірус брязкотіння тютюну — tobacco rattle Tobravirus — може переноситися нематодами або грибами (ще один приклад — вірус некрозу тютюну — tobacco necrpsis Necrovirus).
Переносниками вірусів рослин є членистоногі, нематоди та гриби (рис. 2.). Кожен вірус звичайно має своїх переносників, що належать, як правило, до однієї таксономічної групи. Представники двох класів членистоногих — комахи (Insecta) та павукоподібні (Arachnida) можуть переносити віруси рослин. Безумовний інтерес привертає до себе ряд Homoptera, куди відносяться такі важливі переносники вірусів рослин, як цикади, попелиці та білокрилки.
Представники підряду Aphidinea (попелиці) дуже широко представлені в усьому світі. На території України нараховується близько 77 видів попелиць.
Найважливішими представниками попелиць, що являються переносниками багатьох фітовірусів, є Macrosiphum (Sitobion) avenae (велика злакова), Rhopalosiphum padi (черемхова), Rhopalosiphum maidis (соргова або кукурудзяна), Schizaphis graminum (звичайна злакова), Myzus persicae (зелена персикова), Aphis fabae (бурякова), Macrosiphum euphorbiae (велика картопляна), Myzus (Phorodon) humuli (хмелева), Brachycaudus helichrysi (геліхризова) та ін.
Ротовий апарат попелиці чудово пристосований для інокуляції вірусів. В роботах з застосуванням рослин, що були мічені радіоактивними ізотопами, показано, що при живленні Myzus persicae на рослині-господарі протягом першої години комаха поглинає близько 10 мкл соку рослини, після чого інтенсивність поглинання зростає до 40 мкл/год, що може бути наслідком проникнення стилетів попелиць у флоему — тканину, якій вони надають перевагу. Віруси можуть передаватися попелицями неперсистентно, пересистентно та напівперсистентно. Неперсистентні віруси (потівіруси, карлавіруси, кукумовіруси) передаються при звичайній інокуляції соку уражених рослин і, як правило, викликають симптоми мозаїки. В даному випадку взаємовідносини віруса і переносника не є специфічними. Період порогу передачі займає всього 2 хвилини, а тривалість апробацій, протягом яких віруси активно захоплюються переносником і передаються здоровим рослинам, складає всього 10-30 сек., що дозволяє припускати, що віруси набуваються з клітин епідермісу і вводяться також в клітини епідермісу. Напівперсистентно передаються попелицями представники роду Crinivirus (Closteroviridae). Період порога передачі для вірусу жовтухи буряку складає приблизно 30 хв. Він передається більш ефективно, якщо період доступу продовжується декілька годин. Віруси, що переносяться попелицями персистентно, поділяються на дві групи: ті, що не розмножуються в переноснику (вірус жовтої карликовості ячменю, Luteovirus) та віруси, що розмножуються в переноснику (вірус некротичного пожовтіння салата-латука, Rhabdoviridae).
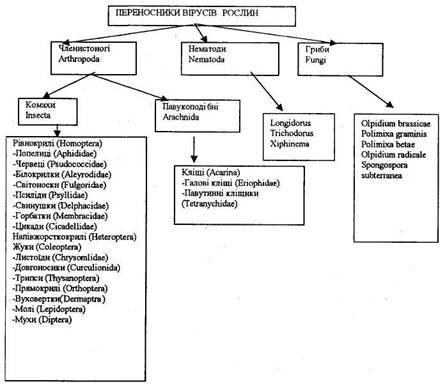
Рис. 2. Переносники вірусів рослин
Близькі види попелиць відрізняються по спроможності передавати вірус. Наприклад, М. persicae, що живиться на хрестоцвітих, уражених одночасно вірусом мозаїки кольорової капусти та вірусом чорної кільцевої плямистості капусти, може передавати обидва віруси, тоді як М. ornatus здатна в цьому випадку передавати лише вірус мозаїки кольорової капусти. На розповсюдження попелиць впливають погодні умови. Розглянемо класичний приклад: у червні 1945 року в Англії була зареєстрована дуже невелика кількість Myzus persicae, але в тей же рік спостерігався великий спалах вірусної інфекції. В червні 1946 року відмічалася найбільша кількість Myzus persicae за останні вісім років, на протязі яких проводилося обстеження, однак рівень ураження в цьому році був низьким. На основі данних, отриманих при роботі з подібними джерелами інфекції, можна припустити, що сезонні коливання частоти ураження такої культури, як цукровий буряк, можливо є наслідком тривалого впливу погодних умов на популяцію попелиці-переносника на протязі більшої частини сезону вегетації. Вони впливають на строки та чисельність попелиць, що мігрують на культуру, на розвиток популяції в середині культури та на її переміщення.
Донедавна білокрилки не відносилися до групи комах, здатних переносити віруси, що можуть викликати епідемії. Говорячи про білокрилок, як правило, припускалося, що ці комахи можуть приводити до економічних збитків в деяких тропічних регіонах та тепличних господарствах, викликаючи пожовтіння листків та призупинення росту рослин. Ще донедавна проблема широкого поширення білокрилок та пов'язаних з ними епіфітотій була актуальною переважно для тропічних та субтропічних регіонів, але вже сьогодні вона постає в агросистемах помірного клімату.
За літературними даними білокрилки можуть переносити представників 4 груп вірусів рослин (родин Geminiviridae, Closteroviridae, Potyviridae та роду Carlavirus).
На сьогодні відомо кілька видів алейродид, здатних переносити віруси культурних рослин. До них належать: білокрилка батата (Bemisia tabaci), білокрилка посрібнених листків (Bemisia argentifolii), оранжерейна білокрилка (Trialeurodes vaporariorum) та облямована білокрилка (Trialeurodes abutilonea).
Фукусі встановив, що вірус карликовості рису передається цикадкою Nepotettix apicalis трансоваріально. Такого роду передача має місце, коли вірус містить самка; коли ж інфекційний тільки самець, передача вірусу неможлива. Фукусі показав також, що цей вірус може також без додаткового інфікування передаватися через яйця на протязі 7 поколінь, що ведуть своє походження від однієї інфікованої самки. На сьогодні є відомості про наявність трансоваріальної передачі у представників Tenuivirus, Reoviridae.
Представники двох родин кліщів класу Arachnida — Tetranychidae та Eriophyidae — можуть переносити віруси рослин. Серед еріофіїд найбільш вивченим клецем-переносником є Aceria tulipae, що здатний передавати вірус смугасатої мозаїки пшениці. Взаємовідносини кліща і віруса вивчені досить детально. Переносник може набувати вірус протягом 15 хв. на хворій рослині і такий же період необхідний для передачі віруса. Після линяння Aceria tulipae не втрачає інфекційності. Передаватися кліщем можуть представники Rhabdo та Potyviridae.
Представники двох основних груп грибів, що уражують корені рослин, виконують роль переносників вірусів рослин — гриб Olpidium spp. серед Chytridiales, та Polymyxa graminis і Spongospora subterranea, що відносяться до Plasmodiophorales. На сьогодні розрізняють два шляхи передачі фітовірусів за допомогою грибів. В першому випадку вірусні частинки, що знаходяться в ґрунтовій воді, прикріплюються на поверхні зооспори гриба. Частинки прикріплюються по всій поверхні зооспори, хоча існує думка, що вірусні частинки, які знаходяться на поверхні джгутиків, втягуються в цитоплазму зооспори, а потім переносяться з протоплазмою в клітину кореня рослини. Таким шляхом передається, наприклад, вірус некрозу тютюну або вірус некрозу огірка грибом Olpidium. Інший шлях передачі спостерігається, наприклад, у віруса моп-топ картоплі та його переносника гриба Spongospora subterranea. Вірус акумулюється грибом, коли він (вірус) розвивається в рослинних клітинах, а не захоплюється з розчинів. Спори у стані спокою залишаються життєздатними і, можливо, зберігають віруси протягом декількох років, а передача вірусів кореням та бульбам здійснюється дводжгутиковими зооспорами.
На сьогодні відомі дві групи вірусів рослин, що можуть передаватися нематодами. Представники групи Tobravirus мають паличкоподібні частки та розповсюджуються нематодами родини Trichodoridae родів Tricodorus та Paratrichodorus. Ізометричні віруси роду Nepovirus, що належить до родини Comoviridae, передаються нематодами родів Xiphinema чи Longidorus родини Longidoridae. У взаємовідносинах переносників з різними вірусами були відмічені лише незначні відмінності. Непо- і тобравіруси нематоди набувають за період від 15 хв. до 1 год і передають їх здоровим рослинам за той же час, при цьому віруси зберігаються в переносниках протягом кількох діб та розповсюджуються як дорослими особинами, так і личинками. Механізм передачі, мабуть, передбачає зворотній зв'язок вірусних частинок зі специфічними поверхнями в шлунковому каналі нематоди. Частки вірусів, що переносяться Longidorus elongatus, зв'язуються з вистилкою стилетної капсули, тоді як віруси, що передаються видами Xiphinema, адсорбуються на стінці стравоходу. Після стадії линяння віруси не зберігаються в переноснику.
Розділ 3. Найбільш поширені вірусні захворювання серед вищих рослин
3.1 Кількісна характеристика та класифікація вірусів рослин
Віруси заражають як вищі, так і нижчі рослини. Відомостей про вірусні хвороби нижчих рослин ще дуже мало. Проте потреба у широких дослідженнях з цього напряму зростає. Дослідження вірусів водоростей і грибів сприятиме розробці біологічних методів боротьби із заростанням (цвітінням) штучних водоймищ, боротьбі з грибними хворобами рослин тощо.
Поки що переважну більшість вірусних хвороб досліджують на вищих рослинах, в основному на зернових, зернобобових та технічних культурах. Це пояснюється великими втратами, які спричинюють вірусні хвороби цим економічно важливим культурам. Вони не менші, ніж від грибних хвороб. Відомо, наприклад, що втрати картоплі від зараження готикою, смугастою та зморшкуватою мозаїкою сягають 50—80 %.
У літературі описано близько 700 вірусних захворювань рослин. Серед збудників цих хвороб добре вивчено понад 150. В Україні зареєстровано понад 115 вірусів рослин. Тривалий час віруси вважались високо специфічними агентами. Тепер уже відомо, що специфічність їх є досить умовною. Наприклад, у літературі наводяться дані про те, що ВТМ уражає не тільки тютюн та деякі інші пасльонові, а й людину, а вірус гепатиту людини — лободу. Ці факти свідчать про відсутність різкої межі між вірусами рослин і тварин за специфічністю. Це має важливе як теоретичне, так і практичне значення, передусім для пізнання властивостей та природи вірусів.
Вірусні хвороби рослин поділяють на дві великі групи — мозаїки і жовтяниці. Мозаїки — хлоротичноплямисті захворювання рослин; концентрація вірусів у їхньому соці досить висока. Віруси мозаїки репродукуються переважно в паренхімних клітинах, передаються механічним способом і комахами. Жовтяниці — вірусні хвороби з характерною великозональною, а інколи загальною некротизацією листків. Концентрація вірусів у соці незначна, локалізація й репродукція вірусів пов'язана з клітинами флоеми.
Найпоширенішим симптомом вірусних хвороб є зміна забарвлення листків та інших зелених органів рослин. На ураженій тканині листка чергуються темно-зелені, світло-зелені або жовті плями. Часто спостерігається хлороз, некротизація і деформація листків та інших органів. Серед мозаїчних хвороб найкраще вивчені звичайна мозаїка тютюну, мозаїки картоплі, буряків, капусти, томатів, злакових і бобових культур.
3.2 Віруси, які вражають певні види рослин
Вірусні хвороби злакових рослин. Великої шкоди завдають вірусні хвороби злаковим культурам. В Україні відомо їх понад 10, а всього в літературі описано близько 60 вірусних хвороб злаків. Найбільшої шкоди зерновому господарству завдають смугаста мозаїка пшениці, мозаїка озимої пшениці, штрихувата мозаїка ячменю, мозаїка кукурудзи тощо. Щороку вони призводять до значних втрат урожаю, а в окремі роки, коли спостерігаються масові епіфітотії, втрачаються тисячі тонн зерна.
Мозаїки картоплі. Велика кількість вірусів зумовлює захворювання цієї цінної культури. До них належать смугаста мозаїка, складчаста мозаїка, крапчаста мозаїка, жовта аукуба-мозаїка, готика картоплі тощо. Подекуди ці хвороби спричиняють виродження високопродуктивних сортів картоплі.
Серед вірусних хвороб, які завдають великих збитків сільському господарству, слід назвати крапчасту мозаїку (її ще називають звичайною мозаїкою) і готику картоплі. Збудником крапчастої мозаїки є Х-вірус картоплі (ХВК) — Solanum virus I (X) Smith. Віріони ХВК ниткоподібні, розмір їх — 500—550 х 10 нм. Часто хвороба проявляється у вигляді ледь помітної світло-зеленої плямистості листків. Збудник легко передається через сік рослин при сумісному зберіганні пророслих уражених і здорових бульб та іншими шляхами. Х-вірус часто бере участь у комплексних захворюваннях картоплі разом з А-, Y-, М- та S-вірусами, посилюючи характерні для них симптоми. В Україні поширена велика кількість штамів ХВК.
Хворобу під назвою веретеноподібність бульб картоплі спричинюють вірусоподібні агенти, названі віріоїдами. За розмірами вони набагато менші відомих нам вірусів і являють собою невеликі молекули РНК; передаються через грунт і з пилком. Про реплікацію цих цікавих вірусоподібних агентів досі ще мало відомо. Є дані про те, що РНК віріоїду веретеноподібності бульб картоплі в заражених клітинах має кільцеву структуру.
Останнім часом поширилась і завдає значної шкоди картоплярству України готика картоплі. Характерною ознакою цієї хвороби є розміщення листків під більш гострим кутом до стебла, що надає рослинам готичного вигляду. Вірус готики передається при різанні бульб ножем, під час щеплень, через бульби, комахами тощо. Найбільш уражується готикою картопля в Сумській, Чернівецькій, Полтавській та південних областях України. Зниження урожайності від цієї хвороби сягає подекуди понад 60 %.
Жовтяниця цукрових буряків. Збудник цієї хвороби (ВЖБЦ) — Beta virus 4 (Roland et Guonjer, Smith) — вперше виявлено в Україні у 1957 р. При ураженні рослин на листках з'являються жовтуваті або жовтогарячі плями невиразних контурів. Пожовтіння починається з верхньої частини листка. На пожовтілому листку утворюються некротичні плями. Пластинка листка потовщується, іноді стає хвилястою. Віріони ВЖЦБ мають ниткоподібну форму. Розміри їх — 250 х х 10 нм. Віруси зимують у коренеплодах маточних буряків. Основним переносником їх є чорна бурякова та зелена персикова попелиці. При ураженні цукрових буряків жовтяницею рослини відстають у розвитку, коренеплоди виростають меншими за масою, значно нижчої цукристості.
3.3 Віруси, які уражують широке коло рослин господарів
Вірус тютюнової мозаїки (ВТМ) Tobacco mosaic virus (TMV)
Вирус тютюнової мозаїки належить до роду Tobamovirus. За морфологією жорсткі палички 300 нм довжиною та 18 нм діаметром. Мають спіральну симетрію з кроком біля 2,3 нм. Геном представлений однією одноланцюговою молекулою +РНК з мол.м. близько 2 х 106 D. Один поліпептид оболонки, мол.м. близько 17 — 18 х103 D. Ефективні імуногени. Більшість членів групи мають помірне коло господарів. Легко передаються механічно, деякі - насінням.
Основним джерелом первинного ураження рослин ВТМ є післяжнивні рослини та їх залишки (стебла, листя, корені). В полі вірус передається соком хворих рослин під час культивації, пасинкування, вершкування, збирання листя та плодів. ВТМ може також переноситись різними комахами з гризучим апаратом гусінню, кониками, слимаками тощо. Нагромаджується і зимує ВТМ в айстрах, капусті, конваліях, подорожнику, маку, хризантемах і багатьох інших багаторічних рослинах, звідки він може переноситись на однорічні рослини.
Крім мозаїки, ВТМ спричиняє на помідорах стрик, внутрішній некроз плодів, нитковидність, папоротевість листя. За деяких екологічних умов (недостатність освітлення, зниження температури, висока вологість) на помідорах з'являються більш тяжкі ураження, які призводять до відмирання ділянок тканин на стеблах та листі. Стрик поширений як в закритому, так і відкритому грунті. Втрати врожаю від хвороби досягають 40-60%. В умовах надмірного азотного живлення стрик проявляється у найбільш шкідливій формі. В останні роки у південних районах країни поширюється внутрішній некроз помідорів, що знижує врожайність до 8-20%. При ранньому ураженні рослини відстають у рості, втрати врожаю перевищують 30-40%. При більш пізньому ураженні мозаїка може бути різко вираженою, але рослини розвиваються, цвітуть і плодоносять. На стиглому листі тютюну ознаки хвороби можуть зникати, хоча рослина буде інфікована вірусом. Втрати врожаю в цих випадках становлять 10-15%. Стійкий сорт тютюну "Імунний- 580.
Рослини - індикатори: N. tabacum (тютюн звичайний), N. sylvestris — системна реакція, Ch. amaranticolor (лобода), N. glutinosa, Phaseolus vulgaris (квасоля звичайна) сорт Пінто, Datura stramonium (дурман звичайний) — локальні некрози.
Циркуляція ВТМ: взимку — зберігання в ґрунті, воді, багаторічних рослинах (подорожник, мак), рослинних залишках, однорічних бур'янах (лобода, дурман), незбираному гарбузинні, коренеплодах буряків, навесні — механічна передача при обробці ґрунту, неспецифічна механічна передача комахами з гризучим апаратом передача між рослинами. Влітку — період найбільшого розмноження, пов'язаний з температурою, вологістю та збільшенням біомаси рослин (картопля, томати, тютюн, перець), співіснування сільськогосподарських рослин та бур'янів, восени — розповсюдження переважно серед бур'янів
Таблиця 1. Рослини, чутливі до ВТМ.
| Beta vulgaris | Буряк |
| Capsicum frutescens | стручковий перець |
| Cucumu sativus | огірок |
| Cucurbitapepo | гарбуз |
| Lycopersicon esculentum | помідор |
| Phaseolus vulgaris | квасоля |
| Solarium tuberosum | картопля |
| Latuca sativa | салат посівний |
| Ch. hybridum | лобода |
| D. stramonium | дурман звичайний |
| Papever rhoeas | мак дикий |
| Physalis floridana | фізаліс |
| Plantago major | подорожник великий |
Безсимптомні рослини:
| Avena sativa | овес посівий |
| Heliantus animus | соняшник |
| Hordeum vulgare | ячмінь звичайний |
| Pisum sativum - | горох посівний |
| Spinacia oleraceae | шпінат городній |
| Triticum aestivum | пшениця м"яка |
| Zea mays | кукурудза звичайна |
| Daucus carota | морква дика |
| Brassica campestris | капуста польова |
| Capsella bursa-pastoris - | грицики звичайні |
| Diantus barbatus | гвоздика бородата |
| Salvia splendens | шавлія блискуча |
| Solarium nigrum | пасльон чорний |
Вірус огіркової мозаїки (ВОМ) Cucumber mosaic virus (CMV)
Вірус огіркової мозаїки належить до родини Bromoviridae, роду Cucumovirus. Віріони ізометричні, можуть бути різних типів, але усі близько 29 нм в діаметрі. Містять 3 геномні і 1 субгеномну (+) РНК.
РНК-1 і РНК-2 містяться в змішаних частках, тоді як РНК-3 і РНК-4 містяться в одній частці. Віріон вміщує 82% білку. Один структурний білок. Молекулярна маса структурного білку 24000D. ТТІ 55 70 °С. Вірус знаходиться у всіх частинах рослини, в цитоплазмі може утворювати ромбоідальні кристали в цитоплазмі.
Природні господарі та симптоми:
Cucumis sativus (огірок посівний) та інпгі представники Cucurbitaсеа — мозаїка, затримка росту; Lycopersicon exculentum (помідор їстівний) — мозаїка, зменшення листової пластинки, затримка росту; Spinacia oleraceae (шпінат городній) — сильні хлорози і затримка росту. Перелік рослин, чутливих до ВОМ, наведено у таблиці 2.
До кола безсимптомних рослин належать: Latuca sativa — салат посівний, Petunia hybrida — петунія гібридна, Spinacia oleracea — шпінат городній, Stellaria media — зірочник середній.
Діагностичні чутливі рослини та симптоми:
Ch. amaranticolor, Ch. quinoa — хлоротичні локальні ураження; Cucumus sativus — системна мозаїка; Vigna unguilata (вігна) — некротичні локальні ушкодження; Lycopersicon sativus — системна реакція (мозаїка, затримка росту).
Таблиця 2.Рослини, чутливі до ВОМ:
| Brassica campestris | капуста |
| Beta vulgaris | буряк звичайний |
| Capsicum annutnn | перець однорічний |
| Coriandrum sativuin | коріанд посівний |
| Cucumus sativus | огірок посівний |
| Cucurbita maxima | гарбуз городній |
| C. pepo | гарбуз звичайний |
| Citrullus lanatus | кавун звичайний |
| Helianthus annuus | соняшник однорічний |
| Lycopersicon esculentum | помідор їстівний |
| N, rustica | махорка |
| N.tabaccum | тютюн |
| Phaseolis vulgaris | квасоля звичайна |
| Pisum sativus | горох посівний |
| Solarium tuberosum | картопля |
| Amaranthus caudatus | щириця хвостата |
| Apium graveolens | селера пахуча |
| Calendula officinalis | календула лікарська |
| Capsela bursa pastoris | грицики звичайні |
| Chenopodium album | лобода біла |
| Ch. hybridum | лобода гібридна |
| Cirsium arvense | осот польовий |
| Datura stramonium | дурман звичайний |
| Gomphrena globosa | гомфрена головчата |
| Lupinus luteus | люпин жовтий |
| L. albus | люпин білий |
| Sonchus oleracea | жовтий осот городній |
| Senacio vulgaris | жовтозілля звичайне |
| Solanum nigrum | пасльон чорний |
| Trifolium repens | конюшина повзуча |
| T. pratense | конюшина лучна |
| Vigna radiata | вігна промениста |
| Medicago sativa | люцерна посівна |
| Lamiun album | кропива глуха |
| Chelidonium majus | чистотіл великий |
| Vicia sativa | горошок посівний |
| Plantago major | подорожник великий |
Для накопичення вірусу використовують рослини Cucumus sativus, N. glutinosa (тютюн клейкий), N. tabacum.
ВОМ передається переносниками неперсистентно, механічною інокуляцією і насінням (з різним ступенем ефективності).
Життєвий цикл Aphids craccivora (люцернова попелиця).
Космополіт. Двудомний вид, мігрує з люцерни на акацію. Засновниці розвиваються з яєць, що зимують на люцерні, дуже рано (на півдні України кінець березня, квітень). Час появи попелиць визначається температурою весняного періоду. Через 8-12 днів попелиці досягають зрілості і починають розмножуватись, в період першого укусу крилаті особини перелітають на інші бобові, акацію, після травня знову мігрує на бобові. Найбільша чисельність відмічається у травні. У червні, в жаркий період літа, попелиць менше, в популяції переважають німфи і крилатки. Комаха веде себе по-різному, в залежності від кліматичних умов
На вторинному господарі в залежності від умов може розвиватися від 2 до 12 поколінь попелиць. Розвиток личинки в залежності від температури складає 5-6 днів при t 24 °С і 21 день при t 10 °С. Оптимальною для розвитку є температура 24 °С .
Максимальна чисельність попелиць на бур'янах та культурних рослинах спостерігається у липні, потім чисельність знижується, і тільки у вересні-жовтні можна спостерігати збільшення розміру колоній.
Житєвий цикл Acyrthosiphon pisum (горохова попелиця).
Попелиця харчується на таких рослинах, як Pisum (горох), Medicago (люцерна), Melilutus (буркун), Vicia (горошок), Faseolus (квасоля), Trifolium (конюшина), іноді — Capsella bursa pastoris (грицики звичайні). Життєвий цикл однодомний. Існує кілька біотипів виду, що віддають перевагу рослинам різних родів. Одна самка дає 150 личинок. За літо розвивається 10 поколінь безкрилих та крилатих незайманок, останні сприяють швидкому заселенні площ. Восени (в залежності від умов) в кінці серпня або вересні — жовтні, з'являється амфогінне покоління, і після запліднення кожна самка відкладає до 10 яєць на люцерні, віці, чині, конюшині. На горох яйця не відкладає, на цю культуру попелиця переходить літом з дикорослих багаторічних рослин в травні — червні. В Середній Європі відмічена зимівля партеногенетичних самок.
Оскільки ВОМ є одним з найменш спеціалізованих вірусів, коло рослин-господарів його надзвичайно широке. Він уражує овочеві культури помідори, перець, огірки, капусту, зернобобові (кукурудзу, горох, квасолю), баштанні (кавун, гарбуз), багаторічні та однорічні рослини. При вивченні рослин резервантів на Україні, ВОМ був виявлений у таких багаторічних рослинах, як осот польовий, подорожник великий. При відмиранні наземної частини згаданих бур'янів, вірус взимку зберігається в їх кореневищах. Наступного року такі рослини є першоджерелом поширення вірусу. Крім багаторічних бур'янів-резервантів, були виявленні й однорічні — дурман звичайний, осот жовтий городній, грицики звичайні. Таким чином, багаторічні бур'яни є місцем перезимівлі вірусу і джерелом інфекції на наступний рік. Однорічні рослини є лише проміжною ланкою в його поширенні.
Відносно стійкі сорти огірків: Донський 175, Молдавський 12. Тираспільський ранній 234, Довгоплідний. Огірки раннього висіву на більш пізніх стадіях розвитку значно менше уражуються вірусом. Особливо велика кількість хворих рослин спостерігається на ділянках, де огірки вирощують як монокультуру. Урожайність знижується від 3,3 % до 80,2 % в залежності від сорту огірків. Висока концентрація вірусу спостерігається при підвищенні температури повітря, зміни світлової інтенсивності; так, короткий день і низька світлова інтенсивність зумовлюють більш тяжкі прояви захворювання і ранню загибель рослин. Висока концентрація вірусу спостерігається при надмірному вмісті в ґрунті азоту та фосфору.
Таким чином, фітовіруси широко розповсюдженні в різних регіонах нашої планети. В умовах України вони мають відповідні шляхи передачі, комах-переносників та коло рослин-господарів, що і зумовлює їх особливості циркуляції в біоценозах та епідеміологічні характеристики.
3.5 Боротьба із вірусними хворобами рослин
Вибір та застосування різних заходів боротьби з вірусними хворобами залежить насамперед від виду хвороби і видів рослин, на які вона поширюється. Загальними для боротьби з вірусними хворобами рослин є такі заходи: 1) виведення і впровадження у виробництво стійких проти вірусів сортів рослин; 2) знищення комах та інших переносників вірусів; 3) ліквідація джерел поширення вірусів у природі (це передусім своєчасне лущення стерні і зяблева оранка, знищення бур'янів на полях, межах, узбіччях доріг, які є джерелом інфекції; дотримання оптимальних строків сівби і проведення противірусного прополювання насіннєвих посівів); 4) використання для сівби здорового безвірусного насінного матеріалу.
Останніми роками серед профілактичних заходів, які запобігають зараженню рослин вірусами, поширення набуває вакцинація рослин (переважно закритого ґрунту) ослабленими штамами вірусів. Інтенсивно проводяться також роботи з оздоровлення рослин методами термотерапії (з використанням спеціальних термокамер), щоб отримати безвірусні клони, хіміотерапії, культур верхівкових меристем тощо.
РОЗДІЛ 4. Біологічний метод: його суть та використання біопрепаратів
4.1 Біологічний метод
Біологічний метод захисту рослин від хвороб полягає у використанні мікроорганізмів-антагоністів та антибіотиків.
Ґрунтові гриби з роду триходерма є активними антагоністами багатьох мікроорганізмів-збудників хвороб. Біопрепарат триходермін, виготовлений на торфі, рекомендується використовувати для обробки насіння 0,3-0,5% суспензією та для замочування коренів розсади томата у суміші з ґрунту, перегною та триходерміну (0,5-1 г триходерміну на рослину). При цьому значно знижується ураженість розсади чорною ніжкою після висадки в грунт.
Застосування компостів у системі біологічного захисту є досить ефективним засобом боротьби з хворобами. Крім цього, вони підвищують мікробіологічну активність ґрунту. Для виготовлення компостів використовують відходи рослинного та тваринного походження. Додавання вермікультури (культури червоного каліфорнійського дощового черв'яка) або каниги (рідини рубця жуйних тварин) до суміші відходів прискорює їхнє розкладання та перегнивання, а також збагачує компости поживними та стимулюючими речовинами. Антибіотик аренарин є ефективним проти бактеріальних хвороб томата (знезараження насіння водним розчином антибіотика в співвідношенні 1:1000).
4.2 Ефективне застосування біопрепаратів при вирощуванні овочевих культур
В умовах сьогодення гостро стоїть проблема пошуку нових перспективних напрямків у сільському господарстві при використанні технологій, безпечних для здоров'я людини, тварин і біоти взагалі. В зв'язку з цим відбувається поступовий перехід від інтенсивного промислового сільськогосподарського виробництва до альтернативного (зокрема біологічного або екологічного), яке передбачає раціональні шляхи використання енергетичних ресурсів і зменшення забруднення навколишнього середовища, одержання високоякісної сільськогосподарської продукції, зберігання і підвищення родючості ґрунту, безвідходне використання сільськогосподарської продукції. Однією зі складових частин екологічного ведення сільського господарства є застосування бактеріальних препаратів, спрямованих на покращення живлення рослин, продукту біоконверсії відходів рослинного походження — біогумату і біологічних засобів захисту рослин. Оскільки овочі є основним джерелом вітамінів, ферментів, мікроелементів, мінеральних солей, інших біологічно активних речовин і входять до незамінних продуктів раціонального харчування людини, що споживаються переважно в свіжому вигляді, вимоги щодо їхньої якості досить високі. Для забезпечення високих кількісних і якісних показників врожайності необхідне дотримання технологій вирощування культури. Застосування біопрепаратів при вирощуванні овочевих культур стимулює ріст і розвиток рослин, покращує азотне та фосфорне живлення, підвищує їхню стійкість до фітопатогенів і, як результат, сприяє підвищенню врожайності та якості продукції, дає змогу не тільки заощаджувати значну кількість енергії, але й створює сприятливий фон для землеробства у цілому, оскільки це сприяє підвищенню родючості грунтів при використанні значно меншої кількості мінеральних добрив і, як наслідок, зниженню рівня забруднення довкілля.
4.3 Бактеріальні препарати і біогумат та їхні властивості
Виготовлення мікробіологічних препаратів і переробка органічних відходів культурою дощового черв'яка належать до екологічно безпечних ресурсозберігаючих технологій.
Розроблена вченими кафедри біотехнології УДХТУ технологія переробки рослинних відходів (соняшникового, гречаного, рисового лушпиння) культурою червоного каліфорнійського черв'яка дозволяє отримати екологічно чисте добриво біогумус, який у водорозчинній формі відомий як біогумат. Біогумат збалансований за вмістом азоту, фосфору, калію та мікроелементів і містить біологічно активні речовини. За останні роки зріс інтерес до розчинних препаратів з біогумусу, які можна використати з більшим виробничим та економічних ефектом завдяки суміщенню з іншими біопрепаратами або новітніми технологіями (крапельне зрошення системи Т-Таре). Застосування біологічних засобів захисту рослин, бактеріальних добрив, біогумусу та водорозчинних екстрактів з нього (біогумату) і є невід'ємною частиною екологічного ведення сільського господарства.
На першій стадії випробовування було вивчено вплив препарату біогумату, який було отримано з соняшникового лушпиння в результаті переробки його каліфорнійським черв'яком родини Lumbricidae, у відповідності з ДСТУ на схожість і проростання насіння помідорів, перцю, капусти, огірків і кукурудзи у лабораторних умовах. Відзначена висока ефективність біогумату від замочування насіння. Методом біотестування було визначено вміст біологічно активних речовин у препараті — 0,28 г/л. Попередньо була визначена й оптимальна доза для замочування насіння — 5 мл на 1 л води.
Агрофіл (ТУ, 565. 10324077003- 92) — біологічний препарат стимулюючих бактерій (Agrubacterium radiobacter 10) використовується для підвищення урожайності овочевих культур. Екологічно чистий. Безпечний для людини та тварин. Застосовується для обробки насіння огірка, томата, перцю, салату та інших овочевих культур. Біопрепарат характеризується такими властивостями: стимулює ріст і розвиток овочевих рослин, підвищує схожість насіння, стійкість рослин до збудників хвороб, покращує мінеральне та водне живлення рослин, що сприяє підвищенню врожайності та прискорює вихід ранньої продукції.
В Південному філіалі Інституту сільськогосподарської мікробіології ведеться робота з розробки технологій одержання та застосування біопрепаратів, основу яких складають мікроорганізми комплексної дії. Новий ефективний штам фосфатмобілізуючих бактерій Enterobacler nimipressuralis 32- 3 виділено з чорнозему південного. Виявлено, що штам утилізує вуглеводи з утворенням органічних кислот, а також продукує лужну фосфатазу, здатен продукувати такі фізіологічно активні сполуки, як індолілоцтову кислоту. На його основі розроблена технологія одержання рідкого біопрепарату ФМБ 32- 3 зі строком зберігання 6 місяців.
Біопрепарат БС11, основою якого є високоефективний антифунгальний бактеріальний штам Paenibacillus polymyxa П, що продукує ріст-регулюючі речовини, має високу швидкість росту і конкурентноздатність у використанні джерел живлення та азотфіксуючу активність. Важливо, що даний штам не проявляє антагоністичної дії по відношенню до колекційних штамів агрономічно корисних мікроорганізмів, на основі яких розробляються біопрепарати. Оскільки штам є продуцентом екзополісахаридів, біопрепарат БСП виготовляється у вигляді гелю зі строком зберігання до 6 місяців і добре прилипає до насіння.
Практична частина
Аналіз способів застосування бактеріальних препаратів і регулятора росту біогумату на овочевих культурах
Біопрепарати використовують для передпосівної обробки насіння. Для розсадних культур рекомендується ще й повторна обробка кореневої системи розсади при висадці її в грунт. Можна використовувати суміш препаратів ФМБ 32- 3 та БСП для інокуляції насіння та обробки розсади. Бактеризація проводиться в день висіву насіння або висадки розсади.
Бактеризація насіння проводиться в день висіву його у вологий грунт. Невеликі партії насіння обробляють вручну під навісом для захисту від потрапляння прямих сонячних променів і перегріву. На чистому брезенті чи поліетиленовій плівці розстилають необхідну кількість непротруєного насіння, біопрепарат розводять водою, ретельно перемішують і рівномірно наносять на його поверхню. Насіння перемішують, почергово піднімаючи протилежні кінці брезенту з метою рівномірного розміщення мікроорганізмів на поверхні насіння. Оброблене насіння згодом відновлює свою сипучість, його затарюють у мішки та висівають.
Механізована обробка насіння. Готується суспензія препарату в воді з розрахунку 1:5. Після інтенсивного перемішування в ємкості необхідного об'єму залити в ємкість протруювальної машини, попередньо відмитої від залишків отрутохімікатів, і обробити насіння перед висівом напівсухим способом з розрахунку 1- 2% суспензії від його маси.
Норми витрати препаратів при передпосівній обробці насіння складають 1- 2% від маси насіння, що обробляється. Наприклад, на 1 кг насіння потрібно 10- 20 мл біопрепарату. При обробці розсади рекомендується поливати кореневу систему рослин розчином препарату 1:100. На одну рослину 100 мл розчину препарату. При сумісному використанні препаратів ФМБ 32- 3 та БСП норма витрати препаратів складає по 1/2 виробничої дози кожною препарату (розчин препарату для поливу кореневої системи (1/2+1/2):100). Обробку кореневої системи розсади можна здійснювати поливною водою (розведення 1:400) через баки розсадосадильної машини.
Обробку сільськогосподарських рослин розчином біогумату (розведення водою 1:200) проводять 2 рази. Першого разу біогуматом обробляють розсаду, другого — поливають рослини через 10- 12 днів після висадки в поле.
Ефективність використання бактеріальних препаратів і регулятора росту біогумату при вирощуванні овочевих культур
Рівень ефективності застосування того чи іншого препарату залежить від реакції виду чи навіть сорту рослин на інокуляцію і від умов вирощування культури. Тому додержання рекомендацій з технології вирощування рослин із застосуванням препаратів є запорукою одержання очікуваних результатів. Враховуючи те, що біопрепарати мають низькі ціни, застосування мікробіологічних препаратів при вирощуванні овочевих культур та зменшення при цьому доз мінеральних добрив веде до зростання економічної ефективності одержання овочів. Незначне зростання затрат дозволяє знизити собівартість продукції, а рівень рентабельності одержання помідорів сорту Шанс підвищується на 6% при обробці біопрепаратом ФМБ 32- 3 і на 8% — препаратом БСП відносно контролю без використання біопрепаратів. Показники економічної ефективності застосування біопрепаратів при вирощуванні капусти білоголової ранньої більш вагомі, що пояснюється особливостями культури, зокрема її одноразовим збиранням. Рівень рентабельності одержання капусти сорту Дітмаршер фрюер при застосуванні біопрепарату ФМБ 32- 3 зростає на 19%, а при обробці БСП — на 35% відносно технології без внесення біопрепаратів.
Застосування бактеріальних препаратів і регулятора росту біогумату на помідорах
На другому етапі проведено польові випробування ефективності біогумату та мікробіологічних препаратів при вирощуванні помідорів протягом 5 років (1998- 2002 рр.) у навчгоспі "Самарський" Дніпропетровського державного аграрного університету.
Застосування біогумату забезпечує прибавку врожаю помідорів 10% до контролю, а сумісне використання біогумату з агрофілом приводить до синергічного ефекту — підвищення врожайності складає 21% (табл. 1).
Таблиця 1. Ефективність застосування біогумату та агрофілу на помідорах сорту Персей
| Варіант | Врожайність, т/га | Приріст урожаю до контролю | |
| т/га | % | ||
| Контроль | 65,47 | - | - |
| Біогумат | 72,14 | 6,67 | 10,2 |
| Біогумат + агрофіл | 79,29 | 13,82 | 21,1 |
| НІР05 | 2,38 | - | - |
Польові випробування біопрепаратів при вирощуванні помідорів сорту Шанс на базі Кримської дослідної станції овочівництва Інституту південного овочівництва та баштанництва і Південного філіалу ІСГМ показали їхню ефективність протягом 2000- 2001 рр. Середній приріст врожаю за 4 роки був найбільшим у досліді під дією біопрепаратів агрофіл і БСП і склав близько 4 т/га, або 14% до контролю. Обробка насіння та розсади помідорів препаратом ФМБ 32- 3 забезпечила дещо нижчу врожайність рослин, яка склала 1,8 т/га, або 6% відносно контролю.
Відомо, що гриби роду триходерма є активними антагоністами багатьох мікроорганізмів-збудників хвороб. Біопрепарат триходермін, виготовлений на торфі, використовували для обробки насіння помідорів 0,3- 0,5% суспензією та для замочування коренів розсади у суміші з грунтом, перегноєм і триходерміном (0,5- 1 г триходерміну на рослину). При цьому розсада краще приживалась завдяки зменшенню ураженості рослин чорною ніжкою після висадки їх у грунт. Вирощування розсади сорту Персей у дослідах з використанням триходерміну і біогумату дозволило підвищити врожайність рослин у середньому за 2 роки відповідно на 8,5 та 15,2% (табл. 2).
Таблиця 2. Ефективність застосуванні біогумату та триходерміну на помідорах сорту Персей
| Варіант | Врожайність, т/га | Приріст урожаю до контролю | ||||
| 1999 | 2000 | т/га | % | |||
| 1999 | 2000 | 1999 | 2000 | |||
| Контроль | 59,76 | 80,95 | ||||
| Триходермін | 65,47 | 86,90 | 5,71 | 5,95 | 9,6 | 7,4 |
| Біогумат | 72,14 | 88,80 | 12,38 | 7,85 | 20,7 | 9,7 |
| НІР05 | 6,07 | 6,66 | ||||
Висновки
Існуючі на сьогоднішній час підходи, як правило, спираються на дослідження та прогнозування розповсюдження збудників вірусних інфекцій рослин, які передаються комахами або нематодами. Вивченню ролі ґрунтів та інших резерватів присвячено зовсім мало робіт. Однак не підлягає сумніву, що вектори можуть первинно інфікуватися не лише з культурних рослин, але і з дикоростучих рослин та ґрунту. При цьому можуть виникати епідемії (епіфітотії)) які фактично викликають повну загибель урожаю на великих площах.
Віруси рослин можуть зберігатися від сезону до сезону в дво- та багаторічних рослинах та в ґрунті, попадаючи туди як прямим (з рослинних залишків або кореневих волосків), так і опосередкованим шляхом (вірусовмісні екскременти); в зв'язку з цим існує загроза зараження нових рослин вірусними інфекціями. Тому необхідно виявляти резервуари вірусних інфекцій в агроценозах, а також досліджувати можливі шляхи розповсюдження вірусів через грунт. Грунт як акумулятор радіонуклідів та важких металів є основною ланкою екосистеми, в якій необхідно вже на ранніх етапах техногенного забруднення оцінювати віддалені наслідки такої дії на біологічну активність ґрунту. Мікроорганізми та віруси, які знаходяться в ґрунті, достатньо швидко реагують на зміну умов оточуючого середовища. З другого боку, дія антропогенного навантаження на ріст та розвиток рослин опосередковано або безпосередньо впливає на фітовіруси. Значною мірою рослинні середовища підвержені руйнівній дії техногенних абіотичних та біозабруднюючих факторів. В екологічно несприятливих регіонах в біоценозах накопичуються в великій кількості фітопатогени, в тому числі і віруси. Коли патоген інтродукується, він може викликати серйозні проблеми для виживання природних членів біоценозу, посилюючи пресинг несприятливих факторів середовища.
Вірусні хвороби рослин поділяють на дві великі групи — мозаїки і жовтяниці. Мозаїки — хлоротичноплямисті захворювання рослин; концентрація вірусів у їхньому соці досить висока. Віруси мозаїки репродукуються переважно в паренхімних клітинах, передаються механічним способом і комахами. Жовтяниці — вірусні хвороби з характерною великозональною, а інколи загальною некротизацією листків. Концентрація вірусів у соці незначна, локалізація й репродукція вірусів пов'язана з клітинами флоеми.
Найпоширенішим симптомом вірусних хвороб є зміна забарвлення листків та інших зелених органів рослин. На ураженій тканині листка чергуються темно-зелені, світло-зелені або жовті плями. Часто спостерігається хлороз, некротизація і деформація листків та інших органів. Серед мозаїчних хвороб найкраще вивчені звичайна мозаїка тютюну, мозаїки картоплі, буряків, капусти, томатів, злакових і бобових культур.
Загальними для боротьби з вірусними хворобами рослин є такі заходи: 1) виведення і впровадження у виробництво стійких проти вірусів сортів рослин; 2) знищення комах та інших переносників вірусів; 3) ліквідація джерел поширення вірусів у природі; 4) використання для сівби здорового безвірусного насінного матеріалу.
Методи виділення з мікроорганізмів речовин, які могли б діяти на патогенні мікроорганізми та були екологічно безпечними, має досить багато переваг в порівняння з іншими препаратами.
Список використаної літератури
1. Актуальные вопросы биологизации защиты растений / Под ред. М.С. Соколова, Е.П. Угрюмова. — Пущино, 2000. — 177 с.
2. Артемьева Н.Н. Методические указания по выявлению и учету вирусных болезней злаков. — М.: Колос, 1971. — 21 с.
3. Атабеков И.Г. Практикум по общей вирусологии. — М.: Из-во Московского университета, 1981. — 191 с.
4. Бабьева И.П., Зенова Г.М. Биология почв. — М.: Изд-во Московского университета, 1983. — 248 с.
5. Білявський Г.О., Падун М.М., Фурдуй Р.С. Основи загальної екології. — К: Либідь, 1993. — 303 с.
6. Биопрепараты в защите растений. Учебное пособие / М.В. Штерншис, Ф.С. Джалилов, И.В. Андреева, О.Г. Томилова. — Мин-во сел. хоз-ва РФ. Ново-сиб. гос. аграр. ун-т, Новосибирск, 2000.— 128 с.
7. Бойко А.Л. Влияние факторов внешней среды на вирусы, инфицируючие растения // Сельскохоз.биология. — 1989. — №5. — С. 120-125.
8. Бойко А.Л. Екология вирусов растений. — К.: Вища школа, 1990. — 167 с.
9. Бондаренко Н. В. Биологическая защита растений. — М.: Агропромиздат, 1987.-278 с.
10. Борьба с вирусными болезнями растений. / Под ред. Х.Кеглер. М.: Агропромиздат. — 1986. — 326 с.
11. Будзанівська І.Г., Поліщук В.П., Тивончук Т.П., Бойко А.Л. Зв'язок між наявністю антигенів фітовірусів в грунті, структурою грунтів та екологічним станом зовнішнього середовища // Вісник аграрної науки. — 1998. — №9. — С 61-63.
12. Вавилов Н.И. Иммунитет растений к инфекционным заболеваниям. - М: Наука, 1986. — 520 с.
13. Вирощування екологічно чистої продукції рослинництва. /Под ред. Дегодюка Е.Г. — К.: Урожай, 1992. — 318 с.
14. Віруси і вірусні хвороби бобових культур на Україні. / Московець СМ., Краев В.Г., Порембська Н.Б. та ін. — К.: Наукова думка 1971.— 136 с.
15. Вірусні та мікоплазмові хвороби польових культур. / Під ред. Ж.П.Шевченко. — К.: Урожай, 1995. — 304 с
16. Вірусні хвороби сільськогосподарських культур/ Московець С.Н., Бобирь А.Д., Глушак Л.Е. / Під ред. Бобиря А.Д. — К.: Урожай, 1975. — С.72-80.
17. Власов Ю. И., Рядко Т.А., Лытаева Г.К. Вирусные болезни овощных культур. — Л.: Колос,1973. — 73с,
18. Власов Ю.И. Вирусные и микоплазменые болезни растений. — М.: Колос, 1992. — 207с.
19. Власов Ю.И., Ларина Э.И. Сельскохозяйственная вирусология. — М.: Колос, 1982. — С. 150-156.
20. Гиббс А., Харрисон Б. Основы вирусологии растений: Пер. с англ. — М.: Мир, 1978. — 430 с.
21. Гнутова Р.В. Серология и иммунохимия вирусов растений. — М.: Наука, 1993. — 300 с.
22. Груздева Л.П., Яекин А.А. Почвоведение с основами геоботаники. — М.: Агропромиздат, 1991. — 448 с.
23. Гулий В.В., Памужак Н.Г. Справочник по защите растений для фермеров. - Кишинев: Universitas, М.: Росагросервис, 1992. — 464 с.
24. Гуральчук Ж.З. Еколого-физиологические аспекты действия цинка на растения // Регуляция минерального питания и продуктивность растений. — К.: Наук, думка, 1991. — С. 102-127.
25. Ижевский С. С, Гулий В. В. Словарь по биологической защите растений. — М.: Россельхозиздат, 1986.— 223 с.
26. Максимов Н.А. Краткий курс физиологии растений, 1958
27. Мусиенко Н.Н., Терневский А.И. Корневое питание растений: Учебное пособие. – К.: Высшая школа, 1989. – 203 с.
28. Мусієнко М.М. Фізіологія рослин. – К.: Либідь, 2005. – 808 с.
29. Нобел П. Физиология растительной клетки. – Л.: Изд-во ленингр. Ун-та, 1983. – 232 с.
30. Твердюков А.П., Никонов П.В., Ющенко Н.П. Биологический метод борьбы с вредителями и болезнями в защищенном грунте. — М.: Колос, 1993. — 160 с.
31. Штерншис М.В. Повышение эффективности микробиологической борьбы с вредными насекомыми. — Новосибирск, 1995. — 194 с.