
Рефераты по авиации и космонавтике
Рефераты по административному праву
Рефераты по безопасности жизнедеятельности
Рефераты по арбитражному процессу
Рефераты по архитектуре
Рефераты по астрономии
Рефераты по банковскому делу
Рефераты по сексологии
Рефераты по информатике программированию
Рефераты по биологии
Рефераты по экономике
Рефераты по москвоведению
Рефераты по экологии
Краткое содержание произведений
Рефераты по физкультуре и спорту
Топики по английскому языку
Рефераты по математике
Рефераты по музыке
Остальные рефераты
Рефераты по биржевому делу
Рефераты по ботанике и сельскому хозяйству
Рефераты по бухгалтерскому учету и аудиту
Рефераты по валютным отношениям
Рефераты по ветеринарии
Рефераты для военной кафедры
Рефераты по географии
Рефераты по геодезии
Рефераты по геологии
Рефераты по геополитике
Рефераты по государству и праву
Рефераты по гражданскому праву и процессу
Рефераты по кредитованию
Рефераты по естествознанию
Рефераты по истории техники
Рефераты по журналистике
Рефераты по зоологии
Рефераты по инвестициям
Рефераты по информатике
Исторические личности
Рефераты по кибернетике
Рефераты по коммуникации и связи
Рефераты по косметологии
Рефераты по криминалистике
Рефераты по криминологии
Рефераты по науке и технике
Рефераты по кулинарии
Рефераты по культурологии
Реферат: Биологические аспекты научения животных
Реферат: Биологические аспекты научения животных
Реферат
Биологические аспекты научения животных
Введение
Механизмы научения животных сложны отчасти потому, что простые формы научения, такие, как привыкание, сосуществуют с более сложными процессами. Например, у людей мы находим привыкание, условные рефлексы, инструментальное научение, рассудочную и когнитивную деятельность. Животные, может быть, не так легко научаются на основе опыта и раccyдочной деятельности, однако многие из них в известной степени несомненно обладают этими способностями. Правда, разные виды животных значительно раздаются по своей экологии и эволюционной истории. Не значит ли это, что естественный отбор формировал способности к научению каждого животного в соответствии с его экологической нишей? Многие биологи поддерживают эту точку зрения. Психологи склонны считать, что некоторые механизмы научения являются общими для многих видов. Оправдан ли такой взгляд? В настоящем рефераты мы пытаемся ответить на этот вопрос.
1. Эволюционные аспекты научения
При научении поведенческий репертуар животного меняется навсегда. Хотя выученное поведение может угаснуть или забыться, животное никогда не вернется к своему прежнему состоянию. Кажущаяся утрата навыка в действительности большей частью представляет собой выработанное торможение предшествующего научения. Научение меняет психологическую структуру животного и тем самым может, по-видимому, изменить его приспособленность.
Процессы научения издавна подвергаются естественному отбору. Поэтому можно ожидать, что последствия научения адаптивны и увеличивают приспособленность животного. Это давно признано и психологами, и этологами, однако систематических попыток рассматривать научение в свете эволюционной теории сделано мало.
При неизменной среде животные ничего не выигрывали бы от научения. Естественный отбор устанавливал бы совокупность простых правил, определяющих надлежащее поведение в той или иной ситуации и входящих в качестве постоянного элемента в психологическую структуру животного. Некоторые свойства среды не меняются, и мы обычно видим, что реакция животных на них стереотипная, врожденная. Так, например, земное притяжение является универсальным свойством среды, и антигравитационные рефлексы имеют тенденцию к устойчивости и стереотипности.
Некоторые свойства среды меняются на основе суточных, лунных или сезонных циклов. Адаптироваться к таким изменениям животному также не обязательно. Фундаментальные черты его физиологической и мотивационной структуры могут меняться периодически в ходе акклиматизации или под действием биологических часов. Обычно животное реагирует на предсказуемые с эволюционной точки зрения изменения среды с помощью заранее запрограммированных изменений своей структуры.
Изменения в течение жизни животного тоже могут быть предсказуемы с эволюционных позиций. В некоторых случаях поведение животного меняется при созревании без участия научения, однако иногда научение для этого необходимо, но является заранее запрограммированным, т. е. происходит на определенной стадии развития животного более или менее независимо от изменчивости индивидуального поведения. Например, у человека дети обладают поразительной способностью к овладению языком в возрасте от двух до семи лет, какой бы язык они не слышали вокруг себя. Подобным же образом у некоторых птиц известны определенные периоды восприимчивости, в которые они обычно научаются пению, слыша своих родителей или сородичей. Такое научение заранее запрограммировано, т. е. происходит в определенный период жизни независимо от индивидуальных условий существования. Ребенок в одном окружении научится немецкому языку, в другом - английскому. Птица в одной среде может научиться одному типу пения, а в другой среде - другому.
Мы увидим, что у молодых животных на определенных стадиях развития происходит импринтинг, т. е. запечатление некоторых свойств среды, в ходе которого они узнают о некоторых особенностях родителей, братьев, сестер или местообитания. Такой импринтинг может повлиять на будущий выбор местообитания, ухаживание и общественное поведение животного. Почему одним животным нужно таким способом узнавать о свойствах их местообитания, родителей или будущих партнеров, тогда как у других видов животных импринтинг неизвестен? Ответить на этот вопрос трудно. Вероятно, в случае выбора местообитания импринтинг помогает животному попасть в условия, сходные с теми, в которых обитают его сородичи, и тем самым увеличить свою приспособленность.
Бейтсон высказал мысль, что некоторым птенцам нужно выделять родителей, которые заботятся о них среди прочих представителей вида, так как родители отличают своих птенцов от чужих, а на чужих могут напасть.
Бейтсон предположил также, что импринтинг, позволяя животному различать близких сородичей, впоследствии помогает ему выбрать несколько отличающегося партнера, за счет чего устанавливается равновесие между инбридингом и аутбридингом. Когда проблемы, встающие перед молодым животным, слегка меняются от поколения к поколению, особи, предрасположенные к узнаванию определенных особенностей среды, например характера местообитания, близких сородичей и источников пищи, могут получить определенное преимущество. Такой вид научения является формой «сопряженного созревания», при котором процессу развития сопутствует приобретение определенного опыта.
Непредсказуемые изменения среды, происходящие в течение жизни отдельной особи, не могут быть предусмотрены наперед запрограммированными формами научения. Для того чтобы адаптироваться к таким изменениям, животное должно полагаться на собственные ресурсы. Способность изменять поведение надлежащим образом перед лицом неожиданных изменений среды обычно считается признаком интеллекта. Но нельзя забывать, что многое можно объяснить обычными механизмами научения.
Исторически сложилось так, что научение исследовалось в искусственных лабораторных условиях, и мало внимания обращалось на специфические потребности и заботы животного. Однако, сомнения в эффективности подкрепления впервые возникли у психологов. Общая реакция на то, что подкрепление не всегда способно изменить поведение так, как намеревался экспериментатор, выразилась в отрицании каких-либо общих законов научения. Была высказана мысль, что способность к научению у разных видов животных строго соответствует экологическим ограничениям, типичным для образа жизни данного вида. Для этологов и эволюционных биологов такая концепция, конечно, заманчива, но означает ли она, что мы обязательно должны отказаться от всяких попыток определить общие принципы научения? Проблема эта противоречива, и прежде чем прийти к какому-нибудь выводу, нужно еще раз обратиться к роли подкрепления в инструментальном научечии.
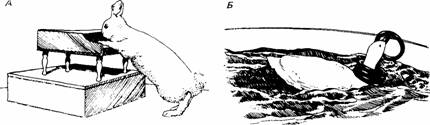
Рис. 1. Примеры фокусов, которым Бреленд обучал животных оперантными условнорефлекторными методами. А. Кролик несколько раз пробегает лапами вверх и вниз по клавиатуре для получения вознаграждения. Б. Утка должна собрать несколько колец, чтобы получим, вознаграждение.
При инструментальном научении животное получает подкрепление за совершение какого-нибудь действия. Подкрепление должно быть произведено в некоторой стимульной ситуации. Определенные черты стимульной ситуации будут регулярно сочетаться с подкреплением, создавая таким образом необходимые условия для классической выработки условного рефлекса. Подкрепление, применяемое в опытах по инструментальному обучению, является также потенциальным классическим БС, который автоматически вызовет врожденную совокупность рефлексов. У кур к этим реакциям относится разрывание почвы лапами в поисках пищи, у свиней - рытье земли рылом - реакции, которые станут классическими условными рефлексами в ответ на стимулы, регулярно сопровождающие еду.
Как же тогда обстоит дело с реакциями, произвольно выбираемыми Скиннером и его последователями для подкрепления?
Мур указал, что почти во всех случаях это не просто случайные реакции, а часть репертуара инстинктивного поведения, обычно связанного с вознаграждением. Так, упомянутое толкательное движение, производимое голубем в скиннеровском «кегельбане», составляет неотъемлемую часть пищевого поведения голубя - отбрасывание земли в сторону для обнаружения семян. Если это толкование правильно, то можно ожидать, что голуби, клюющие ключ для получения пищевого вознаграждения, будут проделывать это иначе, чем за вознаграждение в виде воды. Соответствующий фильм о голубях показывает, что это действительно так. Когда студентов, не знающих, какое из подкреплений было дано, просили различать заснятые опыты с использованием корма или воды, они давали правильный ответ в 87% случаев. Для получения пищи птицы производят сильные, резкие удары по ключу открытым клювом. Когда они клюют, чтобы получить воду, клюв закрыт, а прикосновение к ключу более длительное. При этом часто наблюдаются сосущие движения, характерные для питьевого поведения голубя.
Другие данные относятся к феномену, получившему название "autoshaping". Если многократно сочетать освещение ключа и появление пищи, то голуби начинают клевать ключ без какой-либо описанной выше тренировки. Клевание ключа в большом числе опытов по оперантным условным рефлексам вырабатывается не в результате инструментального подкрепления, а непосредственно по Павлову: с зерном сочетается освещенный ключ, и голуби клюют его, как будто это зерно. Это можно продемонстрировать еще очевиднее, если освещение ключа сочетать с подкреплением, для которого клевание не является естественной реакцией голубей, например с половым вознаграждением. Спариваемые самец и самка помещаются в соседних камерах, разделенных падающей дверцей. Один раз в день включается световой стимул, и скользящая дверца поднимается, чтобы самец мог начать ухаживание. После 5-10 проб самцы начинают выполнять условные реакции на световой стимул. Они направляют свое ухаживание на свет и ведут себя так, как будто это самка.

Рис. 2. Клевание ключа голубями для получения корма и воды: А - для получения воды; Б - для получения корма в опыте типа «autoshaping».
Для того чтобы голубь не получил какого-нибудь инструментального подкрепления в опытах такого типа, можно не давать ему пищу в ответ на клевание ключа. Сила павловского условного рефлекса такова, что голуби, у которых наблюдается "autoshaping", все равно продолжают его клевать. Эти и другие эксперименты, использующие "autoshaping", вне всякого сомнения, установили, что сложные виды поведения - питание, питье и ухаживание - можно получить путем классической выработки условного рефлекса. Они говорят также о том, что многие случаи так называемого оперантного поведения, возникающего при инструментальном научении, на самом деле относятся к классическим условным рефлексам. Но это не значит, что надо обязательно отказаться от закона эффекта.
3. Научение избеганию врагов
Реакции избегания представляют собой форму оборонительного поведения, за счет которого животные сводят к минимуму попадание в ситуации, представляющиеся опасными. Стимулы, вызывающие страх, могут быть сигнальными и вызывать реакцию без какого-либо предварительного опыта. Так же могут действовать и стимулы, с которыми реакция страха или избегания образовала условную связь. К вызывающим врожденное поведение избегания относятся стимулы, связанные с хищниками и с ядовитыми растениями и животными, например змеями и грибами. Врожденное поведение избегания весьма неодинаково у разных видов. Оно выражается в оцепенении, в бегстве, поиске укрытия и тигмотаксисе, в предупреждающих сигналах, служащих для отпугивания нападающего или для оповещения других животных. На рис. 3 и 4 приведены некоторые примеры.
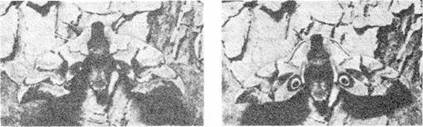
Рис. 3. Отпугивающий узор на крыльях бражника, потревоженного потенциальным хищником. Внезапное появление пятен, похожих на глаза, часто отпугивает мелких птиц.
В естественной среде обитания животное, обучающееся избегать хищников посредством проб и ошибок, пережило бы лишь несколько таких проб. Поэтому животные большей частью обладают врожденными защитными реакциями, рефлекторными по своей природе, хотя и изменяемыми научением. Р. Боллс указал, что экспериментатор в опыте по научению избеганию намеренно или в силу устройства аппаратуры выбирает эффективную реакцию избегания, которая будет или не будет совпадать с врожденной реакцией животного. Степень совместимости врожденного и вырабатываемого в эксперименте поведения сильно влияет на легкость, с какой приобретается реакция избегания.
В течение многих лет психологи сообщали о значительных различиях в легкости, с какой животные научаются реакциям избегания. Так, крысы приблизительно за пять проб научаются перебегать из одного ящика в другой, избегая удара током, но им требуются сотни проб, чтобы для избегания тока научиться нажимать на рычаг. Голуби в этих условиях с большим трудом научаются клевать ключ, но с меньшим - нажимать на педаль. Еще один усложняющий фактор - разница врожденных реакций избегания в зависимости от ситуации. Так, в ответ на удар током крысы обычно убегают, когда есть возможность убежать, и замирают, когда ее нет. Если в такой ситуации присутствует какая-то «мишень», например другой представитель этого же вида, действие тока может вызвать агрессивное поведение. Если ситуация требует, чтобы животное приблизилось к стимулу, связанному с током, тогда выработка реакции избегания становится очень трудной. Такое наблюдается, когда крыса должна нажать на рычаг прямо под источником света, предупреждающим о токе. Если же сигнальный свет находится далеко от рычага, выработать условную реакцию гораздо легче. Применение тока в аппарате с открытым пространством продлевает время, которое крысы проводят в тесном соприкосновении со стенкой. Избегая удара током, эти животные быстрее научаются прыгать на возвышение, находящееся рядом со стенкой, чем на расположенное в середине такого аппарата.
Согласно традиционной теории научения, врожденные оборонительные реакции представляют собой БР. Павловский анализ научения такого типа включает два кардинальных положения. Во-первых, успешные реакции избегания тесно связаны с реакциями, которые обычно вызываются неблагоприятным стимулом. Как мы видели, научение избеганию идет гораздо быстрее, если рефлекс, вырабатываемый экспериментатором, совместим с естественной реакцией животного на применяемое «наказание». Мы вкратце проиллюстрируем этот принцип в другом месте.
Второе Павловское положение гласит, что выработка реакции избегания не должна непосредственно зависеть от возможности избегания как такового. Именно применение отрицательного подкрепления усиливает реакцию, вызываемую этим подкреплением. Отсутствие подкрепления, являющееся или не являющееся результатом избегания, не улучшит процесс классической выработки УР. Здесь, однако, стоит указать, что отсутствие подкрепления иногда не оказывает действия, а иногда оказывает сильное влияние на научение избеганию. Так, например, серебряные караси плывут из одного отделения аквариума в другое, когда это избавляет их от удара током, но они плывут с той же готовностью, когда этот удар неизбежен. И напротив, отключение тока служит эффективным подкреплением, заставляющим морских свинок бегать в колесе или крыс перебегать из одного отделения ящика в другое. Эта методика лучше применяемой при классической выработке УР.
Макинтош указывает, что разница между классическим и инструментальным подходами к выработке реакции избегания может быть связана с вопросом о том, сигнализируют ли внешние стимулы об ударе током или о безопасности и образуют ли реакции на эти стимулы адекватные обратные связи у животного. В тех случаях, когда животное избегает тока, выпрыгивая из ящика или убегая от опасного стимула, само избегание отлично от других форм поведения животного и переносит его в иную внешнюю стимульную ситуацию. Впрочем, для непрерывно плавающего карася уплывание в определенное место не очень отличается от обычного поведения. В то же время для неподвижно сидящей крысы убегание туда, где она не получит удара током, представляет собой особую реакцию с ее очевидными последствиями. Поэтому не следует удивляться тому, что крысе легче научиться реакции избегания такого типа, чем нажиму на рычаг.
Крысы, которые могут нажать на рычаг, чтобы избежать удара током и попасть в отделение ящика, где тока никогда не бывает, научаются нажимам быстрее, чем крысы, которые могут нажимать на рычаг, но при этом не получают доступа в такое отделение. Простой нажим на рычаг оставляет животное в более или менее той же внешней стимульной ситуации, что и раньше, а убегание в другое отделение удаляет его от стимула, связанного с током. Научение избеганию идет гораздо эффективнее в перегороженном ящике с одним «электрическим» отделением, чем в таком же ящике с поочередным включением тока в обеих его половинах, вероятно, потому, что в последнем случае нет участка, ассоциирующегося у крыс с безопасностью.
4. Научение избеганию болезни
Мы видели, что интероцептивные сигналы могут действовать как павловские условные стимулы и что научение играет важную роль в гомеостазе, особенно в регуляции питания. Свидетельства важной роли научения были получены на раннем этапе исследования животных, лишенных определенных ингредиентов пищи. Классические работы Харриса и др. и Рихтера и др. ясно показали, что крысы при нехватке тиамина начинают предпочитать пищу, содержащую это вещество. Скотт и Верни давали испытывающим недостаток тиамина крысам имеющую выраженный вкус пищу с тиамином и безвкусную пищу без тиамина. После того как возникло предпочтение к пище с определенным вкусом, он был перенесен на корм без тиамина, но крысы в выборе еды продолжали ориентироваться на вкус. Это говорит об отсутствии специфическото узнавания тиамина.
Признать, что предпочтение определенной пищи развивается с участием научения, трудно главным образом из-за того, что между пищевым поведением и его подкрепляющими последствиями проходит много времени. В случае недостатка натрия крыса может научиться узнавать, какая пища содержит поваренную соль, попробовав ее на вкус и таким образом получив быстрое подкрепление, стимулирующее выбор того или иного корма. Но при недостатке витамина животное не способно непосредственно обнаружить необходимое вещество в пище. Свойства пищи проявятся только после того, как часть ее будет переварена и в физиологическом состоянии животной; произойдет соответствующее изменение. Хотя высказывалось предположение, что животные научаются связывать стимульные свойства пищи с последующим улучшением самочувствия, эта точка зрения была в то время неприемлема для большинства психологов по двум главным причинам. Во-первых, общее понятие хорошего самочувствия довольно чеопределенно и напоминает концепцию гедонизма. Во-вторых, предполагалось, что промежуток между поеданием пищи и его последствиями гораздо длиннее необходимого для эффективною научения. Были попытки обойти эту вторую трудность постулированием некоего связующею процесса, например продолжительного ощущения вкуса съеденной пищи. Но было показано, что таких возможностей нет.
Гарсия и др. кормили крыс безвредным веществом, а затем вызывали физиологическое последствие независимым от питания способом. После гамма-облучения у крыс возникало отвращение к ранее предпочитавшейся воде с сахарином. Гарсия и др. указали на значение эффекта такого типа для психологии научения животных и привели девять подтверждающих его опытов. Однако прошло еще 10 лет, прежде чем весь смысл этого феномена получил должную оценку и ученые поняли, что потребность в определенном веществе и избегание отравления являются разными сторонами одного и того же явления. Сотни лет люди старались истреблять местные популяции крыс разными средствами, в том числе отравой. Были испробованы самые разнообразные яды, но с ограниченным успехом. Вечной проблемой был вошедший в поговорку страх крыс перед приманками.

Рис. 5. Сравнение классической методики подкрепления и методики его отмены. А. Бег морских свинок в колесе, Б. Условнорефлекторное сгибание ноги у собаки.
Крысы стараются избегать незнакомой пищи и пробовать ее осторожно. Животное, проглотившее небольшой кусочек отравленного корма и выжившее, никогда больше не притронется к нему. Ревуски и Гарсия так описывают лабораторные опыты, демонстрирующие такое поведение:
«Животному дают проглотить пищу с характерным вкусом, например раствор сахарина, а затем подвергают его неблагоприятному последействию, вызываемому такими независимыми от еды средствами, как инъекция яда или рентгеновское облучение. Оправившись, животное будет избегать вещество с этим вкусом. Крыса ведет себя так, как будто думает, что заболела, поев его. Это специфическое отвращение не возникнет, если ухудшение здоровья наступило без предварительного поедания особой пищи или если после ее поедания самочувствие не изменилось. Этот механизм отличается от обычно исследуемых типов научения тем, что может наступить после одного сочетания, даже если между поеданием корма и ухудшением здоровья проходит несколько часов».
Мало что известно о природе внутренних сигналов, с помощью которых животное оценивает свое относительно болезненное или здоровое состояние. Трейсмен предположил, что тошнота и рвота, которые характерны для морской болезни человека и возникают при ненормальной зрительно-вестибулярной стимуляции, происходят потому, что для животного одним из первых показателей отравления является нарушение в вестибулярной системе. Поэтому рвота служит для удаления всякого яда из этой системы, а возникающее вслед за этим угнетенное состояние заставляет животное сохранять покой во время выздоровления. Таким образом, Трейсмен полагает, что у вестибулярной системы, особенно чувствительной и подверженной проникновению мелких молекул, развилась вторая функция - детектора яда. Эта гипотеза еще нуждается в подтверждении.
Болезнь можно считать функцией общего физиологического состояния животного. Можно предположить, что именно степень изменения этой функции животное связывает с пищевыми стимулами. Такой ход рассуждений ведет к следующему выводу:
«Животное не может одновременно научиться предпочитать вещества с определенным вкусом на основе улучшения физиологического состояния и избегать их на противоположной основе. Эти два процесса просто будут гасить друг друга. Например, животное, научаясь избегать определенный вкус из-за следующей за ним болезни, должно также научиться приближаться к источнику этого вкуса после выздоровления».
Розин и Кейлат отмечают, что многие данные, которые, казалось бы, свидетельствуют о положительных предпочтениях, можно истолковать как примеры выработанного отвращения. Проблема состоит в том, что во многих опытах по проверке предпочтений крысы лишены должного выбора. Загорик и Майер позволяли им выбирать между вкусом, связанным с выздоровлением после недостатка какого-либо вещества, вкусом, связанным с таким недостатком, и новым вкусом. Однако крысы не получали корма со знакомым им «безвредным» вкусом. Поэтому возможно, что кажущееся предпочтение пищи, приводящей к выздоровлению, объяснялось просто избеганием крысами остальных видов корма.
В ряде работ сделаны попытки продемонстрировать отчетливое положительное предпочтение, но вопрос этот остается спорным. Положение осложняется тем, что крысы испытывают естественное отвращение к новой пище, хотя у животных, получающих неполноценный корм, эта реакция изменена. Например, крысы, лишенные тиамина, сразу же начинают заметно предпочитать новые виды пищи, даже если в ней нет тиамина, а в старую он добавлен. Розин обнаружил, что, когда крысе, страдающей от отравления или дефицита какого-либо вещества, предоставляется выбор между знакомой безопасной, знакомой опасной и новой пищей, она предпочитаем первый тип корма. По-видимому, крыса научается избегать неполноценного корма, когда в наличии имеется только он. У такой крысы пропадает аппетит, и она отказывается от еды. В результате этого приобретенного отвращения животное сразу же предпочитает новую пищу, что позволяет ей узнать о последствиях ее потребления.
Розин полагает, что крысы делят пищу на четыре класса: новую, знакомую безвредную, знакомую опасную и знакомую полезную. Последняя категория остается спорной. Во всяком случае, обычно принято считать, что крысы очень эффективно и быстро научаются узнавать, что приводит к болезни, и довольно плохо - что способствует выздоровлению. Теперь обратимся к вопросу о том, как происходит это научение.
5. Значимость стимула
Как животное научается связывать потребление определенной пищи с его отдаленными физиологическими последствиями? В интервале между поеданием отравленной пищи и проявлениями токсикоза многочисленные события, с которыми мог бы ассоциироваться токсикоз, в том числе потребление другого корма. Этот вопрос частично можно решить, обратившись к принципу значимости стимула. Согласно этому принципу, сила связи стимула с некоторыми последствиями отчасти зависит от характера этих последствий. Сходными являются понятия готовности и принадлежности. Ревуски и Гарсия считают, что:
«Принцип значимости, определяющий ассоциацию отдаленных физиологических последствий со вкусом еды, состоит в том, что вкус обладает большой ассоциативной силой по отношению к физиологическим последствиям, тогда как ассоциативная сила других экстероцептивных стимулов в этом случае мала. Если же последствием является событие, обычно происходящее во внешней среде, например удар током или получение корма, го наблюдается обратное».
Различные опыты, по-видимому, подтверждают эту точку зрения. Так, например, Гарсия и Кёллинг мет или корм одновременно зрительным, звуковым и вкусовым признаками и сочетали их или с ударом тока или с отравлением. Разным группам крыс предъявляли разные сочетания стимула с последствием. Авторы нашли, что крысы связывали вкус еды с болезненным состоянием, но не с ударом тока, а зрительные и звуковые стимулы - с действием тока, но не с ухудшением самочувствия. Подобным же образом, Гарсия и др. применяли пищевые шарики, покрытые или не покрытые мукой или сахарной пудрой. Схема этих опытов приведена в табл. 1. Крысы поедали шарики разной величины или вкуса. Их «наказывали» электрошоком или облучением. Как и предполагалось, поедание шариков с характерным вкусом заметно подавлялось болезнью, но не ударом тока. Напротив, выбор для поедания шариков различных размеров подавлялся током, но не болезнью.
Некоторые ученые считают, что способность крыс избирательно связывать вкус с болезненным состоянием, наступающим через длительные промежутки времени, представляет собой специализированную систему научения, которая не подчиняется его обычным законам. Однако другие авторы полагают, что почти все свойства обычной лабораторной выработки УР присутствуют и при формировании вкусовых отвращений.
У птиц быстро развивается отвращение к виду пищи. Так, японский перепел в отличие от крыс быстрее научается на основе отравления избегать определенный цвет, а не вкус питьевой воды. Подобные же результаты получены на курах. Если голубей учат нажимать на педаль в присутствии одновременно зрительного и звукового стимулов, то относительное значение их будет зависеть от природы подкрепления. Если им является пища, голуби не отвечают на изолированное предъявление тона, но реагируют на отдельно включаемый свет. И напротив, если подкреплением служит ток, голуби реагируют на тон, а не на свет. Таким образом, очевидно, что во время еды животное обычно обращает внимание на признаки, эффективные с точки зрения выработанного избегания ее физиологических последствий. Крысы в норме реагируют на запах и вкус, а птицы - на внешний вид своей пищи. Если грозит удар током, крысы обращают внимание на зрительные стимулы, а голуби - на слуховые.
Некоторые данные говорят о том, что животные воспринимают удар током как внешний стимул, схожий с нападением соперника или хищника. Для птицы слуховые стимулы, подобные крикам тревоги, могут быть самыми эффективными в ситуациях, где имеется внешняя угроза. Как указывает Макинтош, крысы в течение жизни могут научиться тому, что изменения в зрительной или слуховой стимуляции не связаны с изменениями в их внутреннем состоянии, тогда как изменения вкусовых свойств предсказывают такие изменения. Тенденция взрослых крыс связывать с отравлением скорее вкус, а не зрительные или слуховые стимулы, может быть следствием их способности генерализовать корреляции, с которыми они сталкивались в течение жизни.
Долгий промежуток времени, иногда отделяющий подкрепление от стимула при выработке аверсивной реакции на определенный вкус, ставит более серьезную проблему с точки зрения традиционной теории научения. В обычном опыте по научению подкрепление начинает терять свою эффективность уже после нескольких секунд отставания от стимула, а связь между вкусом и болезненным состоянием может устанавливаться даже при интервале между ними в несколько часов. Высказывалось предположение, что вкус сохраняется долгое время после еды, но ряд исследований исключает такую возможность. Так, например, крысы связывают температуру выпиваемой ими воды с последующим отделенным во времени нездоровьем, а ощущение тепла не может сохраняться дольше нескольких секунд. Кроме того, как мы видели, птицы способны связывать с последующим заболеванием окраску воды. Розин обнаружил, что пернатые научаются избегать определенные концентрации веществ, но трудно представить себе, как такие различия могут распознаваться через полчаса после еды. Некоторые работы показывают, что предъявление второго вкуса в интервале между поеданием незнакомого вещества и возникновением заболевания не нарушает ассоциации первого вещества с нездоровьем. Даже если второй вкус схож с первым, он не будет связываться с плохим самочувствием и не помешает выработке отвращения к предъявленному вначале веществу.
Таблица 1. Схема опыта Гарсии и др. и результаты, ожидаемые на основе значимости стимула
| Переменное СВОЙСТВО | Наказание | Ожидаемый результат |
| Размер шарика | Удар током | Отвращение |
| Размер шарика | Токсикоз | Отвращение отсутствует |
| Вкус | » | Отвращение |
| Вкус | Удар током | Отвращение отсутствует |
Обычно, если крыса отравилась после предъявления незнакомого вкуса, между едой и заболеванием происходит мало событий, которые обладали бы достаточной стимульной значимостью, чтобы ассоциироваться с болезнью. Однако дело обстоит иначе при обычной выработке условной реакции на экстероцептивный раздражитель. Ревуски полагает, что в этих случаях допустимое отставание подкрепления очень непродолжительно, потому что при более длинном промежутке сигналами БР могут стать другие стимулы. Длительные интервалы возможны потому, что только вкусовые стимулы легко образуют связь с болезненным состоянием, а с ударом тока может ассоциироваться множество стимулов. Когда в промежутке между поеданием вещества и последующим заболеванием предъявляется незнакомый вкус, ассоциация с первым веществом нарушается. Предположение Ревуски в какой-то степени примиряет феномены выработки вкусового отвращения с принятой теорией научения, но трудности все же остаются. Так, например, птицы склонны связывать с последующим заболеванием зрительные характеристики пищи, а зрительных стимулов, которые могли бы помешать образованию таких связей, вероятно, довольно много.
6. Перспективы биологического и психологического подходов
Психологов часто обвиняют в том, что они преувеличивают сходство различных животных и слишком полагаются на опыты с лабораторными крысами и голубями. Многие этологи и некоторые психологи считают, что способности определенного вида животных к научению строго соответствуют его экологической нише. Такой взгляд может привести к предположению, что не существует общего процесса научения, присущего многим видам, и что способности данного вида к научению представляют собой комплекс специфических процессов. Так, например, мы видели, что процессы, связанные с выработкой избегания вредной пищи, по-видимому, в некоторых отношениях отличаются от процессов, традиционно изучаемых в лабораториях.
Некоторые типы научения, например научение пению у птиц и импринтинг у молодых животных, совершенно очевидно отличаются от ассоциативною научения. Животное может быть заранее подготовлено эволюцией к тому, чтобы научаться определенным вещам в определенном возрасте.
Другие виды научения, например избегание вредной пищи, тоже считались биологическим приспособлением к образу жизни животного. Такая точка зрения является спорной, потому что при выработке пищевого отвращения можно проследить почти все традиционные механизмы образования УР. Знание тою. какая пища вредна, а какая питательна, какой путь опасен, а какой безопасен, требует некоторой интеграции прогнозных связей между событиями в окружающей среде. Такое знание может быть запрограммировано генетически или приобретено путем научения.
Некоторые психологи, исследующие научение животных, пришли к выводу, что все типы научения в своей основе сходны, поскольку включают общую задачу распознавания причинных связей. Научение позволяет животному связывать причину со следствием и таким образом прогнозировать существенные для себя события. Прогнозные связи основаны на событиях в причинной цепи, свойства которой универсальны. События не происходят без причины и до причины. Мы воспринимаем эти соотношения как сами собой разумеющиеся, и следует предположить, что то же самое свойственно и животным. Лучшими указаниями на будущие события служат их причины, и животные, которые могут обнаруживать и узнавать такие события, будут хорошо приспособлены к взаимодействию с важными всеобщими свойствами природы.
Исходя из этого, некоторые психологи верят в «существование базисного ассоциативного механизма научения, общего для многих видов и предназначенного для обнаружения и хранения информации о причинных связях в окружающей животное среде». Это не отрицает существования способов научения, присущих только определенному виду, или отсутствия у некоторых животных способности к тому или иному виду научения. Просто следует помнить, что у большинства животных один аспект научения общий. Однако такая точка зрения отличается от прежних психологических подходов к научению своей биологической направленностью.